• Структура хроматина меняется с участием комплексов ремоделирования, использующих энергию гидролиза АТФ
• SWI/SNF, RSC и NURF представляют собой большие комплексы, которые характеризуются наличием общих субъединиц
• Сам по себе комплекс ремоделирования не обладает специфичностью по отношению к определенному сайту, однако сборка его должна проходить с участием компонентов транскрипционного аппарата
• Комплексы ремоделирования собираются на промоторах под действием активаторов, специфичных по отношению к определенной нуклеотидной последовательности
• После связывания комплекса ремоделирования с хроматином, фактор может высвобождаться
Геном клетки характеризуется нуклеосомной организацией, однако если промоторная область содержит нуклеосомы, то инициации транскрипции обычно не происходит. При этом гистоны функционируют как репрессоры транскрипции (такое представление известно довольно давно). Процесс активации гена требует изменений в структуре хроматина: существенный момент состоит в том, каким образом аппарат транскрипции получает доступ к ДНК промоторной области.
Способность гена к экспрессии зависит как от локальной структуры хроматина (в промоторном сайте), так и от структуры окружающей области. Структура регулируется за счет местных процессов активации или же изменений, затрагивающих более обширную область хромосомы. К числу наиболее локальных относятся изменения в пределах индивидуального гена, когда изменения в структуре и организации нуклеосом происходят в непосредственной близости от промотора. Более обширные изменения могут затрагивать столь большие регионы, как целая хромосома.
Изменения, затрагивающие большие регионы, контролируют генную экспрессию. Для обозначения зарепрессированного гена в какой-либо области хромосомы используют термин молчащий ген. Под гетерохроматином понимают большие участки хромосом, под микроскопом характеризующиеся более компактной структурой. Молчащие гены и гетерохроматин характеризуются общей особенностью: в обоих случаях с хроматином связаны дополнительные белки, которые прямо или косвенно препятствуют факторам транскрипции и РНК-полимеразе активировать промоторы в определенной области.
Изменения того или иного промотора контролируют транскрипцию определенного гена. Эти изменения могут оказывать или активирующее, или репрессирующее действие.
Динамическая модель транскрипции хроматина основывается на существовании факторов,
способных использовать энергию гидролиза АТФ для вытеснения нуклеосом с участков специфических последовательностей ДНК.
Изменения локальной структуры хроматина представляют собой составную часть процесса контроля генной экспрессии. Гены могут существовать в одном из двух структурных состояний. Гены находятся в «активном» состоянии только в клетках, в которых они экспрессируются. Изменения их структуры предшествуют транскрипции и указывают на то, что ген является «транскрибируемым». Это позволяет предполагать, что приобретение геном «активной» структуры должно быть первым шагом на пути его экспрессии. Активные гены находятся в доменах эухроматина, обладающих повышенной чувствительностью к нуклеазам. Гиперчувствительные сайты создаются на промоторах до момента активации гена.
Между инициацией транскрипции и структурой хроматина существует тесная и постоянная связь. Некоторые активаторы процесса транскрипции непосредственно модифицируют гистоны; в особенности с активацией генов связан процесс их ацетилирования. Напротив, некоторые репрессоры транскрипции деацетилируют гистоны. Таким образом, экспрессия генов контролируется структурным состоянием гистонов поблизости от промотора. Это может представлять собой часть механизма, посредством которого ген поддерживается в активном или неактивном состоянии.
Механизм поддержания локальных участков хроматина в неактивном (молчащем) состоянии связан с механизмом репрессии индивидуального промотора. Белки, ответственные за образование гетерохроматина, действуют на хроматин через гистоны, и модификация гистонов может быть важным этапом его образования.
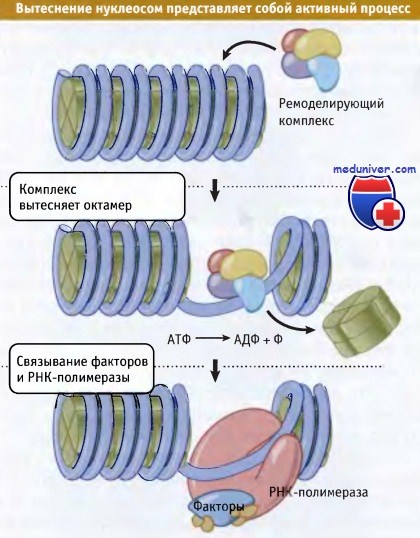
Суммарный процесс реализации изменений структуры хроматина называется ремоделированием хроматина. Он включает в себя механизмы удаления гистонов, которые зависят от поступления энергии. Для удаления гистонов из хроматина необходимо разрушение многих белок-белковых и ДНК-белковых связей. Это не так просто: для диссоциации этих связей требуется энергия. На рисунке ниже представлен принцип динамического ремоделирования хроматина с участием фактора, гидролизующего АТФ. Когда из хроматина высвобождается октамер гистонов, с ДНК могут связываться другие белки (в данном случае факторы транскрипции и РНК-полимераза).
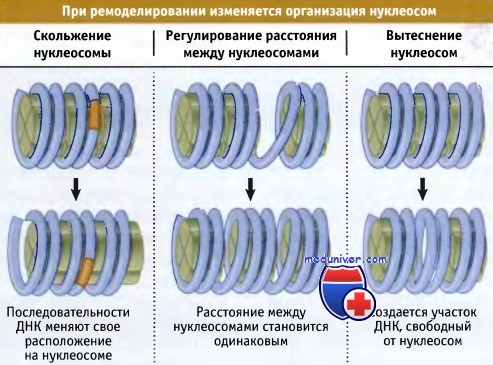
На рисунке ниже суммированы типы изменении, происходящих при ремоделировании хроматина, которые могут быть охарактеризованы in vitro:
• Октамеры гистонов могут продвигаться по ДНК, изменяя соотношение между нуклеиновой кислотой и белком. Это приводит к изменению положения определенных последовательностей ДНК на поверхности нуклеосомы.
• Расстояние между октамерами гистонов может изменяться, что приводит к изменению положения определенных последовательностей по отношению к белку.
• Наиболее радикальные изменения состоят в том, что октамеры гистонов могут полностью удаляться с ДНК, образуя участки, не содержащие нуклеосом. Наиболее часто ремоделирование хроматина происходит при изменении организации нуклеосом в области промотора транскрибируемого гена. Это необходимо для обеспечения доступа аппарата транскрипции к промотору. Однако ремоделирование хроматина также необходимо для обеспечения протекания других процессов, таких как репарация повреждений в ДНК.
Наиболее часто при ремоделировании удаляется один или более октамеров гистонов. Это может быть обнаружено по изменениям характера деградации ДНК хроматина под действием микрококковой нуклеазы: наблюдается утрата сайтов, защищенных от расщепления. Часто при этом возникают сайты, характеризующиеся повышенной чувствительностью к действию ДНКазы I. Иногда наблюдаются менее выраженные изменения, например касающиеся положения отдельной нуклеосомы; это можно определить по утрате лесенки через 10 пн, которая образуется после обработки ДНКазы I. Таким образом, изменения в структуре хроматина варьируют от изменения положения нуклеосом до полного их удаления.
Ремоделирование хроматина осуществляется с участием больших комплексов, которые используют АТФ в качестве источника энергии, необходимой для осуществления этого процесса. Центральным компонентом комплекса является субъединица АТФазы. Обычно комплексы классифицируют в соответствии с типом этой субъединицы — комплексы с близким типом обычно причисляют к одному типу (обычно они содержат также другие общие субъединицы). В таблице на рисунке ниже даны названия этих субъединиц. Два основных типа комплексов представлены SWI/SNF и ISW (ISW заменяет SWI). Дрожжи содержат два комплекса каждого типа. Комплексы обоих типов также находятся у дрозофилы и человека. Каждый тип комплекса может проявлять различную активность при ремоделировании хроматина.
SWI/SNF представляет собой первый из идентифицированных комплексов, участвующих в ремоделировании хроматина. В его названии отражен факт, что многие субъединицы кодируются генами, идентифицированными в экспериментах на клетках 5. cerevisiae с помощью мутаций SWI и SNF. Эти мутации генетически взаимодействуют с мутациями в генах, кодирующих компоненты хроматина. Это гены SIN1 и SIN2, кодирующие негистоновый белок и гистон Н3 соответственно. Гены SWI и SNF требуются для экспрессии различных индивидуальных локусов (включая -120 генов или 2% генома S. cerevisiae). Для экспрессии этих локусов может оказаться необходимым, чтобы комплекс SWI/SNF обеспечил ремоделирование хроматина в области соответствующих промоторов.
Ремоделирующий комплекс может вызывать скольжение нуклеосом вдоль ДНК,
вытеснять нуклеосомы или изменять расстояние между ними.
В экспериментах in vitro SWI/SNF проявляет каталитическое действие, и в дрожжевой клетке содержится только 150 копий комплексов. Все гены, кодирующие субъединицы SWI/SNF, не являются абсолютно необходимыми, что позволяет предполагать, что у дрожжей должны существовать другие пути ремоделирования хроматина. Комплекс RSC представлен большим количеством копий и, вероятно, является более необходимым. Он действует на -700 локусов-мишеней.
Комплексы SWI/SNF могут ремоделировать хроматин in vitro или без потери всех гистонов, или с удалением их октамеров. Для обоих процессов характерно общее промежуточное состояние, при котором структура нуклеосомы — мишени меняется, приводя или к повторному образованию (ремоделированию) нуклеосомы на той же ДНК, или к перемещению октамера гистонов на другую молекулу ДНК. Комплекс SWI/SNF меняет чувствительность нуклеосомной ДНК к ДНКазе I и изменяет ДНК-белковые контакты, которые сохраняются после ее высвобождения из нуклеосом. Субъединица SWI2 представляет собой АТФазу, которая обеспечивает энергией процесс ремоделирования, осуществляемый с помощью SWI/SNF.
Между ДНК и октамером гистонов существует много точек связывания — по результатам исследования кристаллической структуры их насчитывается 14. Для того чтобы октамер сместился на новую позицию или же полностью потерял связь с ДНК, должны разорваться связи во всех точках контакта ДНК с белком. Каким образом это происходит? Некоторые механизмы можно исключить, поскольку мы знаем, что при ремоделировании однонитевая ДНК не образуется (и комплексы не обладают хеликазной активностью). Согласно современному представлению, комплексы, относящиеся к классам SWI и ISW, используют энергию гидролиза АТФ для раскручивания ДНК на поверхности нуклеосомы. Некоторые косвенные данные свидетельствуют о том, что при этом создается механическое усилие, которое позволяет небольшому участку ДНК отсоединиться от поверхности и занять новую позицию.
Одним из важных процессов, катализируемых комплексами ремоделирования, является скольжение нуклеосом. Сначала оказалось, что комплекс группы ISW изменяет положение нуклеосомы, но не вызывает отрыва коровой частицы от ДНК. Это достигается за счет скольжения, при котором октамер перемещается вдоль ДНК. Удаление N-терминального участка гистона Н4 предотвращает скольжение, однако мы достоверно не знаем, каким образом эта часть молекулы гистона участвует в перемещении. Комплексы SWI/SNF обладают такими же свойствами; процесс блокируется при создании барьера на ДНК. Это позволяет считать, что мы имеем дело с реакцией скольжения, при которой октамер гистонов более или менее непрерывно движется вдоль ДНК, не теряя с ней контакта.
Одна из загадок функционирования комплекса SWI/SNF заключается в его размере. Комплекс состоит из 11 субъединиц и обладает суммарной молекулярной массой 2 х 106. Он должен мешать функционированию РНК-полимеразы и нуклеосом. Кроме того, непонятно, каким образом все компоненты комплекса взаимодействуют с ДНК, которая удерживается на поверхности нуклеосомы. Однако обнаружен транскрипционный комплекс, обладающий полной активностью, который называется РНК-полимераза II холофермент. Он содержит саму РНК-полимеразу, все TFII факторы, за исключением TBP и TFIIA, и комплекс SWI/SNF, который связан с CTD концевым участком молекулы полимеразы. Фактически препараты холофермента могут содержать весь комплекс SWI/SNF. Это позволяет предполагать, что ремоделирование хроматина и узнавание промоторов являются скоординированными процессами и осуществляются с участием одного комплекса.
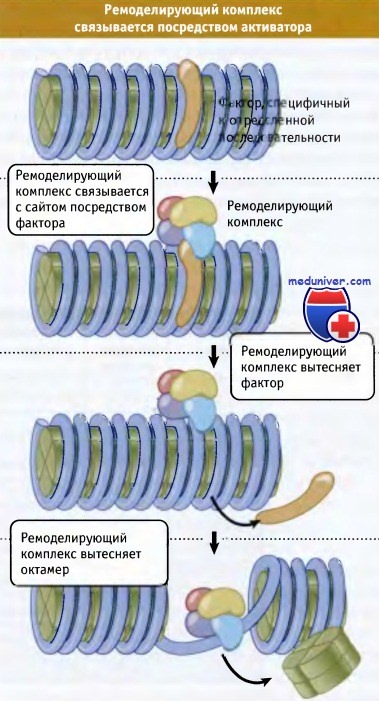
Каким образом комплексы присоединяются к определенным сайтам хроматина при ремоделировании? Сами по себе они не содержат субъединиц, связывающихся со специфическими последовательностями в ДНК. Поэтому предложена модель, согласно которой функционирование комплексов зависит от активаторов или (иногда) от репрессоров. Модель функционирует по механизму «hit and run», при котором после связывания комплекса ремоделирования активатор или репрессор удаляется.
Ремоделирование хроматина достигается за счет изменений в состоянии гистонов, в особенности модифицирования N-терминальных участков гистонов Н3 и Н4. Эти участки включают 20 N-терминальных аминокислот и отходят от нуклеосомы, располагаясь между витками ДНК. Они могут быть модифицированы по нескольким сайтам за счет метилирования, ацетилирования или фосфорилирования. При модификации гистонов создаются сайты связывания для негистоновых белков, в результате чего изменяются свойства хроматина.
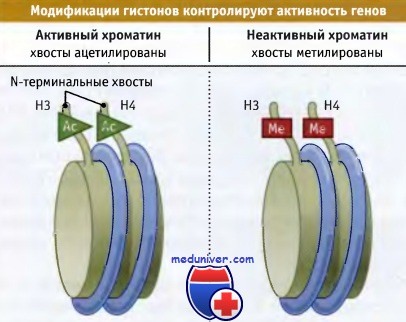
Количество нуклеосом, белки которых подвергаются модификации, различно. Модификация может быть локальной и, например, ограничиваться нуклеосомами, расположенными в области промотора. Она может представлять собой более общий процесс и затрагивать хромосому целиком. Рисунок ниже иллюстрирует положение, согласно которому ацетилирование связано с активацией хроматина, а метилирование вызывает его инактивацию. Впрочем, это простое правило выполняется не всегда, и важную роль играет характер модифицируемого сайта, а также сочетание различных модификаций. Поэтому известны исключения, когда, например, в активном хроматине находятся гистоны, метилированные в определенных положениях.
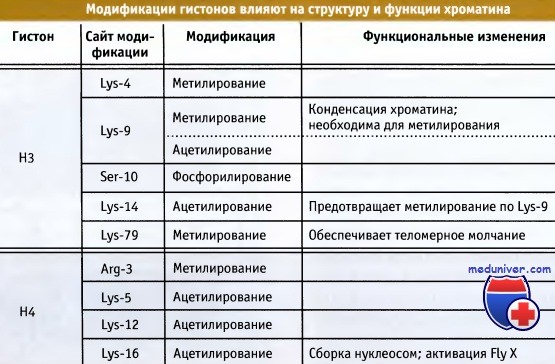
О специфичности модификаций свидетельствуют данные о том, что для многих ферментов на отдельных гистонах существуют специфические сайты. В таблице на рисунке ниже перечислены некоторые модификации. Большинство сайтов подвергается только одному типу модификации. В некоторых случаях модификация одного сайта может оказывать активирующее или ингибирующее действие на модификацию другого. Представление о том, что комбинация сигналов может быть использована для определения типа хроматина, иногда называется гистоновый код.
Ремоделирующие комплексы подразделяются на типы в зависимости от природы АТФазных субъединиц.
Ремоделирующий комплекс связывается с хроматином посредством активатора (или репрессора).
Ацетилирование гистонов Н3 и Н4 характерно для активного хроматина,
в то время как метилирование для неактивного.
Большинство сайтов в гистонах подвержены одному, определенному типу модификации,
однако некоторые из них могут модифицироваться разными ферментами.
С некоторыми из этих модификаций могут быть связаны определенные функциональные изменения.