Регуляция pH клетки и внеклеточной среды через обмен натрия
• pH внутриклеточной среды и объем клетки поддерживаются за счет процессов Na+/Н+-обмена.
• В результате действия переносчика Na+/HCO3- снижается закисление среды за счет прямого транспорта в клетку ионов HCO3-.
Как отмечалось в предыдущем разделе, многие транспортные белки используют энергию трансмембранного градиента Na+ для перемещения ионов и других метаболитов через плазматическую мембрану.
В данной статье мы рассмотрим два других переносчика, Na+/Н+-обменник и Na+/HCO3--котранспортер, которые используют энергию трансмембранного градиента Na+. Эти транспортеры принимают участие в регуляции pH, важного фактора, определяющего функциональную активность белков, поскольку свойства многих из них зависят от величины pH.
Nа+/Н+-обменники и Nа+/НСO3--котранспортеры помогают поддерживать необходимый кислотно-щелочной баланс внутри и вне клетки. Наиболее важной буферной системой во внеклеточной среде, например в плазме, является обменная система бикарбонат/двуокись углерода, которая функционирует при участии фермента карбоангидразы.
Обычно кровь обладает слабощелочным значением pH (7,4), которое находится под эффективным контролем, поскольку функциональная активность многих клеток и белков существенным образом зависит от величины pH.
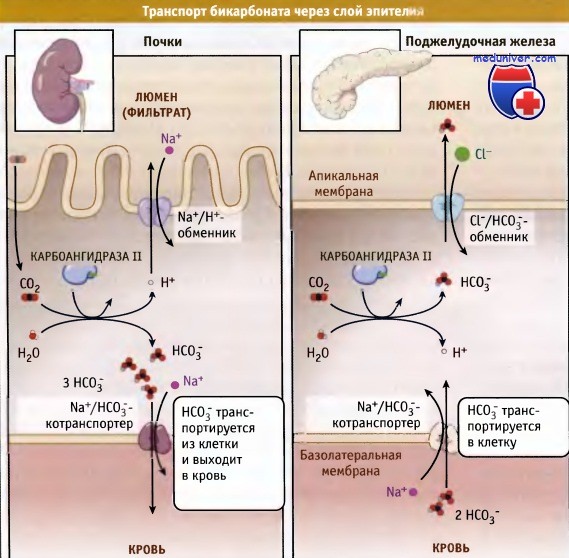
Модели, описывающие транспорт бикарбоната через эпителиальные клетки в почках и поджелудочной железе.
Легкие и почки относятся к органам, принимающим участие в поддержании кислотно-щелочного равновесия плазмы за счет процессов экскреции летучих (СO2) и нелетучих кислот. Почки контролируют pH крови за счет регуляции транспорта НСO3- через эпителиальные клетки, выстилающие начало проксимального канальца.
В этих клетках существуют два источника СO2: диффузия через мембрану и синтез с участием специфических ферментов. В проксимальных канальцах карбоангидраза катализирует образование Н2O и СO2 из НСO3- и Н+. Двуокись углерода свободно диффундирует в эпителиальные клетки проксимальных канальцев, где происходит синтез НСO3- и Н+ из Н2O и СO2 с участием карбоангидразы. Таким образом, секреция протонов (кислоты) в почечный фильтрат (мочу) за счет Nа+2/Н+-обменника апикальной мембраны (NHE) сочетается с транспортом в кровь равного количества эквивалентов бикарбоната через Na+/НСO3-котранспортер, находящийся в базолатеральной мембране проксимального канальца.
Такой сопряженный выход из клетки ионов Н+ и НСO3- не сопровождается изменениями электрического заряда и представляет собой важный механизм выделения в мочу нелетучих кислых эквивалентов; он служит важным звеном гомеостаза. Суммарный эффект заключается в реабсорбции бикарбоната из фильтрата плазмы в виде СO2, который затем, как НСO3-, транспортируется в плазму через апикальную мембрану.
В то же время NHE в проксимальном канальце способствует реабсорбции ионов Na+ из плазматического фильтрата, после чего они транспортируются назад, в плазму. Ключевым элементом, регулирующим кислотность и реабсорбцию в проксимальном канальце, является Na+/К+-АТФаза.
Суммируя вышесказанное отметим, что экскреция Н+ и вход в клетку Na+ через апикальную мембрану происходят с участием специфических симпортеров и антипортеров.
В отличие от почек эпителиальные клетки поджелудочной железы и других секреторных органов секретируют бикарбонат через апикальную мембрану. В клетках секреторных органов за счет градиента ионов Na+ происходит котранспорт НСO3- в стехиометрическом отношении 1 Na+ на 1 или более НСO3-.
В базолатеральной мембране клеток поджелудочной железы котранспортер Na+/HCO3- обеспечивает захват бикарбоната из плазмы, НСO3- образуется под действием карбоан-гидразы, и он транспортируется в желудочно-кишечный тракт посредством Cl-/НСO3--обменника в апикальной мембране.
Панкреато-дуоденальная секреция бикарбоната играет важную роль в буферизации кислотной среды, создающейся за счет желудка, а также в активации важнейших пищеварительных ферментов.
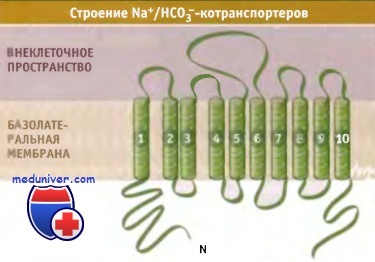
В почках и поджелудочной железе экспрессируются различные изоформы Na+/HCO3--котранспортеров. На рисунке ниже представлена модель, описывающая их строение и предложенная на основании анализа первичной структуры белков.
Na+/Н+-обменник представляет собой интегральный мембранный белок плазматической мембраны и внутриклеточных органелл. Наряду с ролью, которую он играет в поддержании внутриклеточного pH, этот обменник участвует в регуляции объема клетки, осуществляет системный контроль за электролитами, метаболизмом кислот и гомеостазом жидкостей организма. Na+/Н+-обменник катализирует транспорт ионов Na+ и Н+ в направлении их концентрационных градиентов. Он является вторичным активным переносчиком.
Градиент ионов натрия, направленный в клетку и образованный за счет функционирования Na+/К+-АТФазы плазматической мембраны, обеспечивает продвижение протонов в обратном направлении со стехиометрией 1 Na+ на 1 Н+. Поток ионов натрия через мембрану регулирует клеточный объем и поглощение солей и воды через эпителиальные клетки.
Регуляция функционирования Nа+/Н+-обменников осуществляется при изменении внутриклеточного pH и под действием различных внешних стимулов, например ростовых факторов, гормонов, а также механических стимулов, например осмотического стресса и распластывания клеток.
В физиологических условиях в результате деятельности Nа+/Н+-обменника плазматической мембраны удаляется избыток кислот, накопленный за счет метаболизма и за счет их поступления извне. Вместе с бикарбонат-транспортной системой Nа+/Н+-обменники играют критическую роль в поддержании кислотно-щелочного равновесия в цитоплазме.
Изоформа 1 Nа+/Н+-обменника (NHE1) рассматривается как прототип обменника плазматической мембраны клеток млекопитающих; другие изоформы отличаются уровнем тканевой экспрессии и характером локализации в мембране.
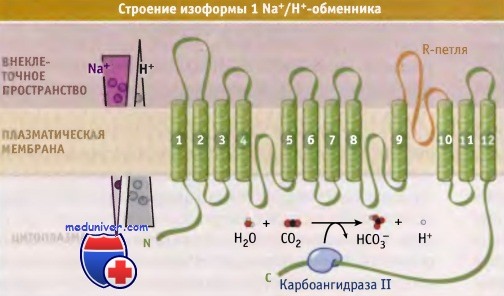
Все изоформы Nа+/Н+-обменника имеют общее строение. Компьютерное моделирование вторичной структуры позволило предположить, что изоформы состоят из 12 встроенных в мембрану и сходно расположенных N-концевых сегментов с консервативной структурой и из более вариабельных по структуре С-концевых областей, содержащих многочисленные сайты фосфорилирования и способных связывать регуляторные белки и медиаторы. Предполагается, что N-терминальный трансмембранный домен содержит каталитическую коровую структуру, принимающую участие в обмене Na+/H+.
Большая петля, частично расположенная в мембране, а частично вне клетки (R-петля), между трансмембранными сегментами 9 и 10, напоминает структуру поровой петли К+-каналов и, вероятно, является составным элементом транспортного пути ионов. Два Na+/H+ белковых обменника могут образовывать гомодимерную структуру.
Бактерии обладают Nа+/Н+-белками-обменниками, которые для выведения Na+ в обмен на поток Н+ в клетку используют энергию электрохимического градиента Н+ по сторонам мембраны.
В клетках Escherichia coli основным Nа+/Н+-обменником является NhaA, который регулирует содержание в клетке ионов Na+ и значение pH и обеспечивает выживаемость бактерий в условиях засолености или щелочности окружающей среды. NhaA существует в цитоплазматической мембране в виде димера, и каждый мономер состоит из 12 трансмембранных сегментов.
Предполагаемое строение изоформы 1 Na+/Н+-обменника (NHE1).
Функция NHE1 регулируется несколькими путями. Показано связывание карбоангидразы II, которое приводит к стимуляции транспортной активности NHE1.
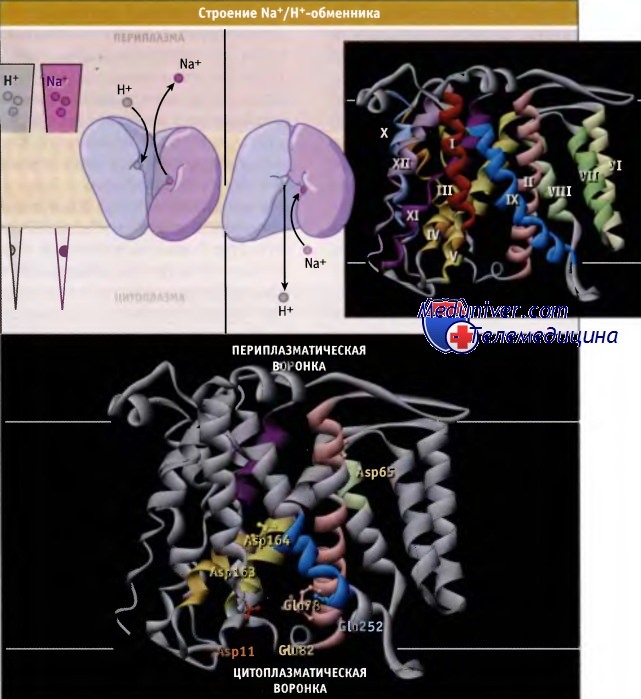
Слева показано направление Na+- и Н+-градиента по сторонам плазматической мембраны клеток животных.
На рисунке ниже представлена кристаллическая структура мономера NhaA в неактивной конформации, которая характерна для кислых значений pH. По обеим сторонам мембраны белок имеет воронкообразные поверхности, которые выстилаются отрицательно заряженными остатками аминокислот, притягивающими катионы.
В центре мембраны расположены аминокислоты Asp163 и Asp164, образующие сайт связывания Na+. При кислых значениях pH только Asp164 оказывается на пути потока ионов, и транспорт в цитоплазму блокируется спиралью Х1р.
Вероятно, что транспорт Na+ и Н+ с участием NhaA происходит по механизму попеременного доступа. Согласно предложенной модели, изменение pH цитоплазмы в щелочную сторону индуцирует изменения конформации спирали IX, что, в свою очередь, вызывает реориентацию спиралей ХIр и IVc.
При этом открываются сайты связывания Na+ и снимается цитоплазматический барьер. Связывание Na+ с цитоплазматической стороны приводит к небольшим дополнительным изменениям конформации спиралей ХIр и IVc, закрытию обменника со стороны цитоплазмы, и к выходу Na+ в периплазму.
После высвобождения Na+ с периплазматической (внеклеточной) стороны должно произойти протонирование Asp163 и Asp164, что приводит к конформационным изменениям, в результате которых сайт связывания Na+ открывается со стороны цитоплазмы, где должно происходить депротонирование остатков аспартата.
Изменение активности Nа+/Н+-обменника в клетках человека является причиной развития некоторых заболеваний, таких как гипертония, диарея, диабет, а также тканевых повреждений, вызванных ишемией (за счет прекращения или замедления кровотока).
Развитие ишемии мозга или миокарда приводит к увеличению интенсивности Na+/Н+-обмена. Это вызывает повышение концентрации внутриклеточного Na+ и вторичное увеличение содержания внутриклеточного Са2+, что обусловлено активацией Na+/Са2+-обменника в обратном режиме.
Перегрузка клетки Са2+ вызывает каскад процессов, которые приводят к сердечной аритмии, или к изменениям нервной ткани после инсульта. Ингибирование активности Na+/Н+-обменника препаратами может оказаться благоприятным при ишемии или после нее и широко исследуется в качестве подхода к лечению некоторых заболеваний.
Схематическое строение и модель рентген-анализа структуры бактериального Na+/Н+-обменника, NhaA, объясняющие транслокацию Na+/H+ и механизм регуляции pH.
Кристаллическая структура представлена белками. Остатки, изменения в которых влияют на регуляцию pH и транспорт катионов, показаны вместе с боковыми цепями.
Схематическая модель транслокации, функционирующая по принципу попеременного доступа, построена на основании структурных данных.
Показаны градиенты Na+ и Н+ по сторонам мембраны. Модель построена на основании данных Protein Data Bank file 1ZCD.