• Ran представляет собой небольшую ГТФазу, которая присутствует в ядре и цитоплазме всех эукариотических клеток
• Комплекс Ran-GAP способствует гидролизу ГТФ под действием Ran, а комплекс Ran-GEF участвует в обмене ГДФ на ГТФ на Ran
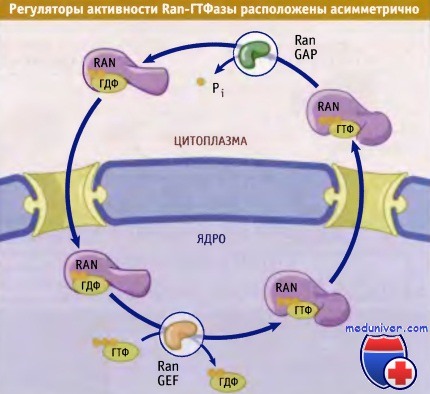
• Ran-GAP цитоплазматического происхождения, в то время как Ran-GEF находится в ядре
• Ran контролирует ядерный транспорт за счет ассоциации с кариоферинами и влияя на их способность связываться с карго
В каждый момент времени в клетке множество белков транспортируются в ядро и выходят из него. Перемещение белков происходит при взаимодействии NLS или NES белков карго с белками семейства кариоферинов. Переносчики мигрируют в обоих направлениях, связывая карго в одном компартменте и высвобождая его по другую сторону ЯПК. Каким образом контролируется направление транспорта? Ответить на этот вопрос помогает исследование свойств небольшого мономерного белка Ran, связывающего ГТФ. Этот белок ассоциирует с транспортерами и мигрирует между ядром и цитоплазмой. В белке Ran нет сигналов NLS и NES.
Как и другие белки, связывающие ГТФ, Ran способен очень медленно гидролизовать ГТФ, образуя ГДФ. Существуют белки-регуляторы, стимулирующие ферментативную активность Ran. К их числу относятся два белка, Ran-GAP и Ran-GEF. Первый представляет собой белок, активирующий ГТФазу Ran и приводит к быстрому гидролизу связанного ГТФ с образованием формы Ran, находящейся в комплексе с ГДФ. Ran-GEF представляет собой фактор, обменивающий гуаниловый нуклеотид, и присоединяет к Ran новые молекулы ГТФ. Этот белок обеспечивает удаление связанного ГДФ и замещение его на ГТФ.
Таким образом, в зависимости от регулятора, с которым он взаимодействует, Ran находится в форме, связанной либо с ГТФ, либо с ГДФ. Ran-GEF (который также обозначается как Reel) и Ran также частвмот в сборке митотического веретена.
В цитоплазме Ran существует в форме, связанной с ГДФ, а в ядре фермент связан с ГТФ.
Это обусловлено характером распределения ферментов, влияющих на его способность связываться с ГТФ и гидролизовать нуклеотид.
Для простоты остальные белки, участвующие в транспорте, не показаны.
Ключ к пониманию того, каким образом обеспечивается направленность ядерного транспорта, лежит в различной локализации Ran-GAP и Ran-GEF. Как показано на рисунке ниже, Ran-GAP локализован в цитоплазме, и некоторое количество комплекса связано с филаментами на цитоплазматической стороне ЯПК. Это означает, что в цитоплазме Ran находится в форме, связанной с ГДФ, поскольку Ran-GAP будет стимулировать гидролиз связанного ГТФ. Напротив, Ran-GEF локализован в ядре. Таким образом, в ядре Ran должен быть в ГТФ-связанной форме, поскольку после импорта Ran-ГДФ в ядро Ran-GEF может заменить связанный с Ran-ГДФ на ГТФ. Это асимметричное распределение эффекторов Ran предполагает, что концентрация Ran-ГДФ должна быть выше в цитоплазме, а концентрация Ran-ГТФ — в ядре. Такое предположение было подтверждено изящным экспериментом, схема которого представлена на рисунке ниже.
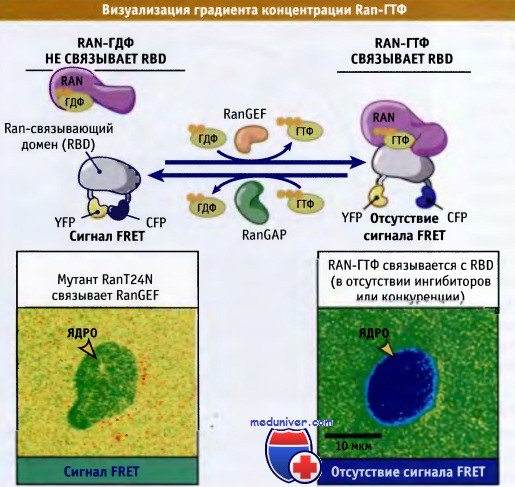
Из белка, который связывался с Ran-ГТФ, но не с Ran-ГДФ, и двух флуоресцирующих белков получали белковый гибрид. Флуоресцирующие белки включали желтый (YFP) и голубой белки (CFP). Затем в клетке вызывали экспрессию белкового гибрида. Когда этот белок не связан с Ran-ГТФ, его флуоресцирующие домены располагаются рядом, и между ними происходит взаимодействие. Это взаимодействие называется флуоресцентным резонансным переносом энергии (FRET), и его можно наблюдать с помощью флуоресцентного микроскопа. Таким образом, FRET будет наблюдаться, когда белковый гибрид находится в присутствии Ran-ГДФ. Напротив, если присутствует Ran-ГТФ, то белковый гибрид с ним связывается. Это связывание предотвращает взаимодействие доменов YFP и CFP, и FRET не наблюдается. На микрофотографии, представленной на рисунке ниже справа внизу, видно присутствие в ядре голубого сигнала FRET, что свидетельствует о наличии больших количеств Ran-ГТФ именно в этой органелле, а не в цитоплазме.
Слева представлены результаты контрольного эксперимента, выполненного на клетках, экспрессирующих мутантную форму Ran (RanT24N), которая связывается с Ran-GEF и препятствует замещению ГДФ связанного с Ran дикого типа на ГТФ. Эти клетки характеризуются пониженным содержанием Ran-ГТФ в ядре, что проявляется наличием слабого зеленого сигнала FRET. Эти результаты показывают, что Ran преимущественно связан с ГТФ в ядре клетки, и с ГДФ — в цитоплазме. Установлено, что концентрация Ran-ГТФ в ядре примерно в 200 раз превышает его концентрацию в цитоплазме.
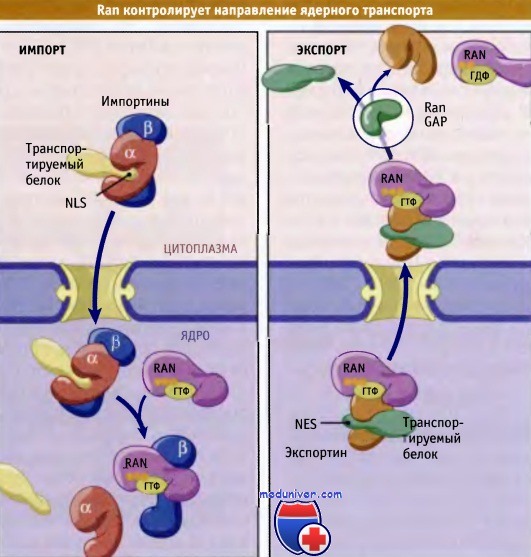
Ran контролирует направление ядерного транспорта, взаимодействуя с кариоферинами и влияя на их способность связываться с карго. Эффект Ran зависит от того, связывается он с импортером или с экспортером транспортируемого белка. Рассмотрим импортен, рецептор ядерного импорта. В цитоплазме он связывается с белком карго, содержащим NLS. Попав в ядро, комплекс импортин-карго связывается с Ran-ГТФ, что приводит к его перегруппировке и к отщеплению карго Таким образом, комплексы импортин-карго, находящиеся в цитоплазме, устойчивы, но диссоциируют после связывания в ядре с Ran-ГТФ. Отметим, что, хотя концентрация Ran-ГДФ в цитоплазме высока, он не является необходимым ни для связывания импортина с карго, ни для осуществления процесса импорта.
Напротив, для связывания экспортинов с белками карго, содержащими NES, необходим комплекс Ran-ГТФ. В присутствии Ran-GEF в ядре поддерживается высокий уровень Ran-ГТФ. Ran-ГТФ, экспортин и карго кооперативно связываются, образуя трехмерный комплекс. После того как этот комплекс через пору вышел в цитоплазму, он вступает в контакт с Ran-GAP, который гидролизует ГТФ. При этом тримерный комплекс диссоциирует, и белок карго высвобождается.
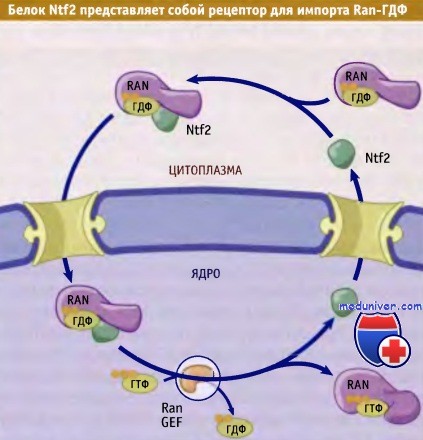
Критическую роль в процессах ядерного транспорта играет распределение Ran, связанного с ГТФ и ГДФ в ядре и цитоплазме. В большинстве случаев Ran концентрируется в ядре. Однако из ядра вместе с экспортином постоянно выходит некоторое количество Ran-ГТФ. После гидролиза ГТФ и высвобождения карго, Ran-ГДФ поступает назад в ядро при участии собственного транспортного рецептора, который называется Ntf2. В ядре, под действием Ran-GEF, связанный с Ran ГДФ замещается на ГТФ, что приводит к отщеплению Ntf2 от Ran. После этого Ntf2 выходит из ядра и принимает участие в повторном цикле импорта Ran-ГДФ.
Наряду с Ran-GAP и Ran-GEF, существуют другие белки, которые связывают Ran и изменяют его активность. Один из таких белков, RanBP1 находится на поверхности ЯПК, обращенной в сторону цитоплазмы. Этот белок стимулирует ГТФазную активность Ran при участии Ran-GAP, увеличивая эффективность высвобождения карго. Второй белок — RanBP3, находится в ядре, где обеспечивает взаимодействие экспортина с ЯПК. Для того чтобы создать высокую концентрацию Ran на ЯПК, некоторые нуклеопорины связывают Ran с ядерной и цитоплазматической стороны. Один из них, Nup 358, также называется RanBP2.
Ran также выполняет важную роль в митозе, и за счет взаимодействия с импортином b регулирует доставку ключевых белков к митотическому веретену.
С помощью Ran-связывающего домена (RBD), способного связываться только с Ran-ГТФ,
можно определить относительные концентрации Ran-ГТФ и Ran-ГДФ, характерные для различных клеточных компартментов.
Импортины связывают белки в цитоплазме и высвобождают их в ядре после ассоциации с Ran-ГТФ.
Напротив, с Ran-ГТФ в ядре образуются комплексы для экспорта.
Экспортируемый белок высвобождается в цитоплазму при гидролизе ГТФ.
Траспорт Ran в ядро происходит с участием небольшого белка, который называется Ntf2.