Функции Rab-белков и Rab-ГТФазы в адресовании белков
• Так же как и мономерные ГТФазы из семейства Sar/ARF участвуют в образовании белков окаймления транспортных везикул, семейство Rab-ГТФаз участвует в адресовании этих везикул в мембраны-реципиенты
• На каждом этапе везикулярного транспорта участвуют различные представители семейства Rab
• Белки, которые отбираются или активируются системой Rab. называются даунстрим-эффекторами. Они включают такие удерживающие белки, как длинные фибриллы и большие мультипротеиновые комплексы
• С помощью удерживающих белков везикулы присоединяются к мембранным компартментам, а компартменты друг к другу
Когда произошел отбор белков карго, и они включились в транспортную везикулу, должно произойти ее слияние с мембраной следующего на пути компартмента. Это не простая задача, принимая во внимание существующее богатство выбора внутриклеточных мембран. Клетка располагает по крайней мере двумя независимыми группами белков, Ras-подобными ГТФазами, которые называются Rabs, и удерживающими белками. Эти белки должны обеспечивать точность процесса адресования до момента слияния мембран.
ГТФазы, относящиеся к семейству Ras, представляют собой первые из идентифицированных белков, участвующих в адресовании. Они были обнаружены в ходе отбора мутантов, при изучении секреции белков у дрожжей. Например, для транспорта из ЭПР в аппарат Гольджи требуется белок Ypt1p, а для транспорта из аппарата Гольджи в плазматическую мембрану — белок Sec4p.
В отсутствие функционально полноценного белка Sec4p, транспортные везикулы, образующиеся в аппарате Гольджи, не могут сливаться с плазматической мембраной. Гомологи этих белков в клетках млекопитающих были обнаружены при скрининге библиотек кДНК головного мозга крысы и отсюда известны под названием белки Rab («rat brain»).
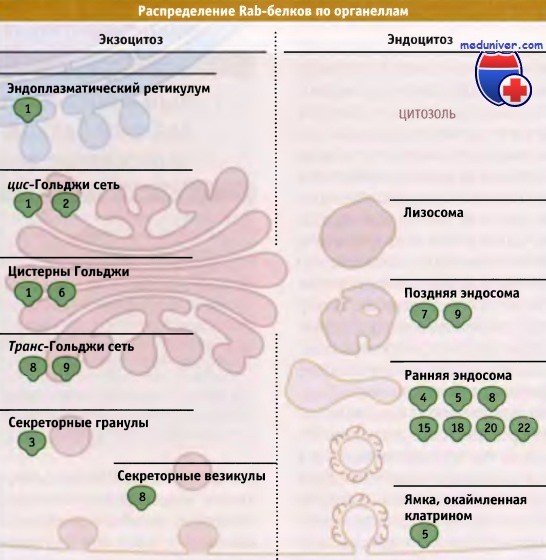
Локализация некоторых Rab-белков млекопитающих на органеллах.
В настоящее время в почкующихся дрожжах обнаружено около двенадцати, а в клетках млекопитающих около 60 белков, относящихся к семейству Rab/Ypt. Учитывая количество отдельных этапов транспорта при эндоцитозе и на секреторном пути, существует, вероятно, достаточное количество Rab-белков для того, чтобы за каждый отдельный этап отвечал по крайней мере один белок. В общем, Rab-белки локализуются в тех органеллах, где они функционируют.
Так же как и другие мономерные ГТФазы, белки Rab/Ypt могут быть в неактивной, связанной с ГДФ, и активной, связанной с ГТФ форме. Белок Rab, связанный с ГДФ, находится в цитозоле, а форма его, связанная с ГТФ, ассоциирована с мембранами. Так же как и белки Ras, каждый белок семейства Rab/Ypt содержит каталитический домен ГТФазы и липидный якорь, расположенный на С-терминальном участке молекулы. Якорь состоит из двух прениловых (геранил-гераниловых) цепей, содержащих по 20 углеродных атомов, и помогает белку закрепиться на мембране. Последовательность, расположенная на С-терминальном конце, определяет компартмент (или компартменты), с которым связывается Rab-белок.
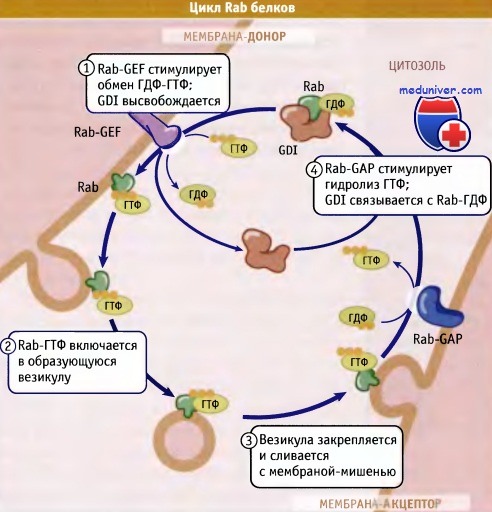
Ассоциация Rab-белков с мембранами представляет собой регулируемый процесс. На рисунке ниже представлена одна из моделей, предложенных для описания локализации белков Rab/Ypt. Белок Rab в форме, связанной с ГДФ, присутствует в цитозоле в виде комплекса с ингибитором отщепления гуаниловых нуклеотидов (GDI). При блокировании функции прениловой группы, которая способствует ассоциации с мембраной, GDI препятствует образованию мембранно-связанной формы Rab, переводя ее в растворимое состояние.
На подходящей мембране-доноре Rab-ГДФ освобождается от GDL и происходит замена ГДФ на ГТФ под действием мембранно-связанного GEF. Однако остается неизвестным, каким образом Rab-белок узнает мембрану. Возможно, что роль «рецепторов» Rab выполняют связанные с мембраной GEF. Так или иначе, активированный Rab-ГТФ включается в образующуюся везикулу.
Rab-ГТФ (и удерживающие белки) позиционируют везикулу на акцепторной мембране. После этого под действием активирующего белка, обладающего ГТФаз-ной активностью и специфичного для Rab (Rab-GAP), происходит гидролиз ГТФ. Образующийся Rab-ГДФ затем связывается с GDI и возвращается в исходный компартмент.
С использованием методов генетики и биохимии было обнаружено много эффекторов Rab, представляющих собой белки, которые преимущественно связываются с ГТФ-содержащей формой Rab и выполняют свои дальнейшие функции. Одна группа таких эффекторов представлена белками, имеющими форму спирализован-ных фибрилл, которые выполняют роль удерживающих белков и образуют начальную связь между транспортной везикулой и мембраной-мишенью. В данном разделе описываются удерживающие белки, связывающие транспортные везикулы с мембранами аппарата Гольджи.
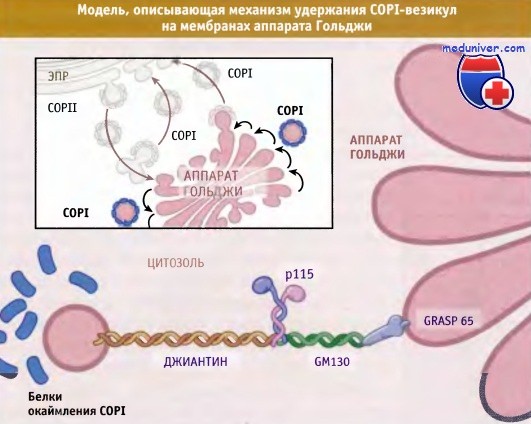
Белок Rab1 представляет собой гомолог дрожжевого белка Ypt1p у млекопитающих и участвует в транспортных процессах из ЭПР в аппарат Гольджи, а также в самом аппарате Гольджи. Rab1-ГТФ связывается с p115, биспиральным белком, являющимся эффектором Rab. Одна из функций белка p115 состоит в удержании везикул COPI на мембранах аппарата Гольджи. Эту же функцию также выполняют еще два биспиральных белка, джиантин и GM130. В экспериментах с использованием рекомбинантных белков in vitro впервые было продемонстрировано взаимодействие джиантина и GM130 с Rabl.
Как показано на рисунке ниже, джиантин закрепляется на мембране COPI-везикул с помощью домена, пронизывающего мембрану, в то время как GM130 фиксируется на мембранах Гольджи с помощью миристилиро-ванного белка GRASP65. Rab1 связывается с джиантином, GM130 и p115, однако роль этого связывания в процессе удержания везикулы остается неясной. Очевидно, что комплекс, образующийся под действием Rab1, удерживает COPI-везикулу на мембране Гольджи, хотя остается непонятным, важно ли это удержание для прямого или обратного транспорта.
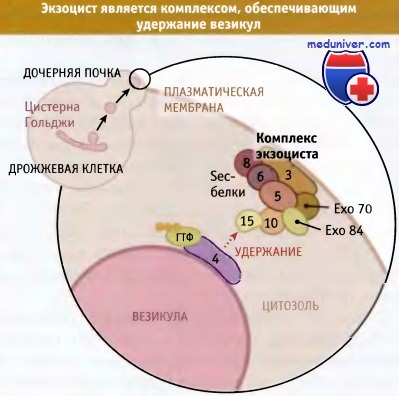
Наряду с удерживающими белками, обладающими фибриллярной структурой, существует семейство больших мультипротеиновых комплексов, которые также участвуют в ранних стадиях адресования везикул. К числу их относится комплекс TRAPP, расположенный с входной стороны аппарата Гольджи, комплекс COG, участвующий в транспортных процессах внутри органеллы, и комплекс экзоцист, расположенный на плазматической мембране.
У дрожжей экзоцист представляет собой комплекс, состоящий из семи белков, который способствует взаимодействию везикул, прошедших аппарат Гольджи, с плазматической мембраной на конце растущего дочернего пузырька.
В клетках эпителия млекопитающих гомолог экзоциста находится в месте контакта апикального и базолатерального доменов. Нарушение функции экзоциста за счет мутации или при введении антител приводит к замедлению или полному блокированию секреции белков. По-видимому, этот эффект является следствием нарушения способности транспортных секреторных везикул позиционироваться на местах последующего слияния с плазматической мембраной.
Модель, описывающая цикл Rab-белков между цитозолем и мембранами
(другие белки, участвующие в перемещении, не показаны).
COPI-везикулы удерживаются на мембранах Гольджи спирализованными белками.
В данной модели движение COPI-везикул ограничивается этими белками, которые помогают выбрать мембраны цистерн, межу которыми продвигаются везикулы.
Rab1 связывается с GM130, р115 и с джиантином, однако точное строение комплексов неизвестно и поэтому не показано.
Экзоцист представляет собой комплекс белков цитозоля,
которые обеспечивают удержание секреторных везикул на плазматической мембране.