Протеинкиназы регулирующие вступление клетки в митоз
• Во многих эукариотических клетках основной контрольной точкой служит переход от G2 в М-период цикла
• С переходом G2-M связана активация нескольких протеинкиназ
G2-фаза клеточного цикла связана с подготовкой клетки к митозу. В течение этой фазы большинство клеток растет, и, таким образом, после деления у них сохраняется постоянное ядерно-цитоплазматическое отношение. Во время этой фазы, т. е. до момента вступления клетки в митоз и начала сегрегации хромосом, также узнаются и корректируются ошибки репликации ДНК. Как клетки начинают митоз, когда завершились все процессы? В настоящем разделе мы рассмотрим роль протеинкиназ во вступлении клетки в митоз.
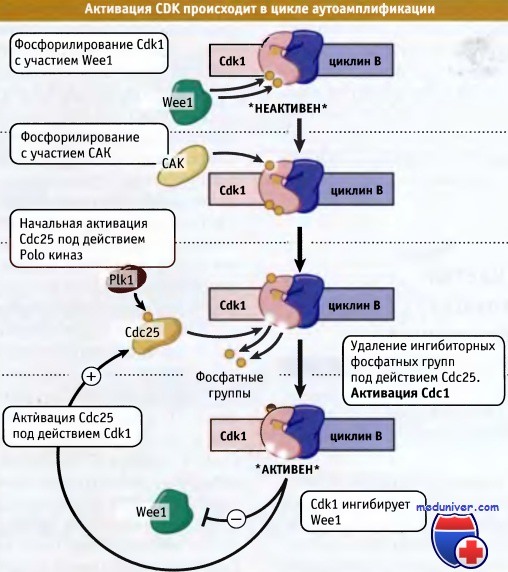
Основной киназой митоза, способствующей переходу G2-M, является комплекс Cdk1-циклин В. По мере накопления митотических циклинов, они связываются с Cdk1. Комплекс накапливается, однако он находится в неактивном состоянии, поскольку белок фосфорилирован при участии представителя семейства киназ Weel. Если в клетках отсутствует активность Weel, то Cdk1 не подавляется. При этом вступающие в митоз клетки достигают небольшой величины, т. е. являются wee. Киназа Mik1 является гомологом Weel и участвует в фосфорилировании и ингибировании активности Cdk1 при пролонгировании S-фазы. Гомолог Weel, Myt1, локализован исключительно в эндоплазматическом ретикулуме (ЭР), в то время как другие представители этого семейства находятся в клеточном ядре. Эти киназы фосфорилируют тирозиновый (Tyr15) или расположенный рядом треони-новый остаток (Thrl4), что приводит к ингибированию активности фермента.
Дефосфорилирование остатков Tyr15 или Thr14 контролирует активацию Cdk1 и вход в митоз. Этот процесс осуществляется Cdc25 фосфатазой. У делящихся дрожжей ген cdc25, в отличие от weel, является необходимым. Если фосфатные группы не удаляются, то митоз не наступает Для гарантии в клетках млекопитающих присутствуют три разных изоформы Cdc25, которые обеспечивают наступление этого ключевого процесса.
В клетках высших эукариот наиболее подробно исследован один из этапов активации Cdk1, протекающий с участием Cdc25. В этих клетках активация Cdc25 происходит с участием Polo-киназ (PLKs), представителей семейства, отличающегося от CDK. После активации Cdc25 с участием PLK и последующего увеличения активности Cdk1 дальнейшая активация Cdc25 происходит под действием самой Cdk1. Такая форма положительной обратной связи при активации Cdk1 приводит к сильному увеличению активности киназы, что приводит клетку к вступлению в митоз. Активность Weel также отрицательно регулируется за счет фосфорилирования Cdk1 с тем чтобы обеспечить резкую активацию Cdk1, необходимую для вступления клетки в митоз. На рисунке ниже схематически представлены процессы активации Cdk1.
У многоклеточных эукариот вхождение клетки в митоз и сам митоз находятся под контролем двух различных CDK-циклиновых комплексов. Например, в клетках млекопитающих Cdk1-циклин А регулирует некоторые процессы в митозе, включающие конденсацию хромосом и их выстраивание у митотического веретена. Варианты циклина В, обладающие разной локализацией и периодически меняющие активность в цикле, вероятно, регулируют другие процессы митоза за счет фосфорилирования различных субстратов. В результате многолетних исследований было найдено много субстратов Cdk1, некоторые из них являются реальными кандидатами на роль эффекторов Cdk1 в митозе.
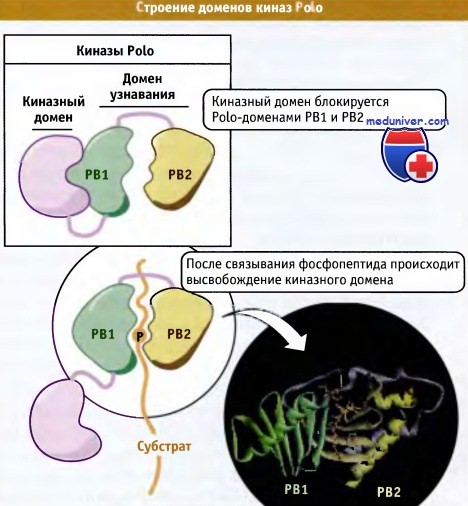
Хотя Cdk1 считается основным регулятором митоза, другие представители семейства киназ также играют важную роль в различных митотических процессах, и в настоящем разделе мы рассмотрим три группы таких киназ. К их числу относится уже упомянутое выше семейство PLK. Впервые эти киназы были обнаружены в мутантах Drosophila, у которых были нарушены некоторые процессы митоза, а в дальнейшем PLK были найдены в клетках многих эукариот, где они выполняют роль пускового фактора митоза, участвуют в созревании центросом, образовании веретена, сегрегации хромосом и в цитокинезе. В то время как геном Drosophila и дрожжей кодирует только одну киназу, в клетках позвоночных образуется до четырех разных представителей семейства PLK. При этом наиболее функционально близким к ферменту дрожжей и Drosophila является Plk 1. Все ферменты содержат N-концевой киназный домен и С-концевой участок, включающий одну или несколько консервативных последовательностей, которые называются доменами Polo (или Polo boxes). Строение концевых участков и активация PLK показаны на рисунке ниже.
Фосфорилирование Cdk1 подготавливает фермент к активации, однако он поддерживается в неактивном состоянии.
Polo- киназы (Plks) активируют фосфатазу Cdc25, которая затем активирует небольшое количество Cdk1, удаляя фосфатные группы.
После активации эта киназа фосфорилирует Cdc25, что приводит к увеличению уровня активного фермента.
Наряду с этим, при активации некоторого количества Cdc1 она фосфорилирует и инактивирует Weel.
Этот цикл аутоамплификации приводит к резкой активации Cdc1.
Домены Polo выполняют роль внутриклеточных функциональных доменов и участвуют в позиционировании киназ на многочисленных сайтах причаливания, расположенных на центросомах, кинетохорах (белковые структуры, связывающие хромосомы с концами микротрубочек), митотическом веретене и цитокинетическом кольце. Эти домены связываются с мотивом фосфо-Сер/Трео-Про, который представляет собой фосфорилированную консенсусную последовательность в Cdk1 и в других киназах, обладающих сродством к пролин-содержащим сайтам, таких как МАР киназы. Эти данные позволяют предполагать, что PLK присоединяются к белкам, которые предварительно были фосфорилированы Cdk1 или другими киназами, хотя возможно, что in vivo сайты причаливания PLK получают дополнительную информацию от присоединяемых компонентов. Такой механизм мог бы обеспечить координацию активности различных митотических киназ, а также совместную регуляцию Plk1 и других протеинкиназ. Он также помог бы объяснить, почему в митозе PLK активируются параллельно или вслед за Cdkl и другими киназами. По-видимому, в различных сайтах для PLK существует множество субстратов. Все известные субстраты принимают участие в формировании веретена и в цитокинезе.
Еще одна группа киназ, участвующих в митозе, представлена семейством NimA-подобных киназ (NEK). Эти ферменты были обнаружены у A. nidulans. История открытия NEK хорошо иллюстрирует тезис о том, насколько важно использовать различные организмы для исследования процессов, контролирующих клеточный цикл. Каждый из объектов исследования обладает своими особенностями, а в совокупности они обеспечивают получение разнообразной ценной информации. Мутанты nimA организма A nidulans заблокированы в С2-периоде, хотя активность Cdk1 в клетках высока. Это говорит о том, что самой по себе активации Cdk1 еще недостаточно для наступления всех событий митоза. Действительно, оказалось, что активность NIMA в митозе меняется параллельно с изменением активности Cdk1. После секвенирования первой обнаруженной киназы гомологичные NEK были найдены в клетках многих эукариот, и было продеменстрировано их участие в нескольких событиях митоза, включая конденсацию хромосом и расхождение центросом.
Третья группа киназ, которая привлекла внимание исследователей в связи с участием в процессах митоза, представлена семейством Aurora. Так же как Cdkl и PLK, киназы Aurora участвуют во многих процессах, включая конденсацию и сегрегацию хромосом, функционирование кинетохора, созревание центросом, образование веретена и цитокинез. Киназы Aurora впервые были обнаружены в почкующихся дрожжах, где они представлены одной киназой. Однако впоследствии было показано их присутствие в клетках многоклеточных эукариот и человека, где они принадлежат к одной из трех следующих групп: Aurora А, В, или С.
Все ферменты семейства Aurora имеют общую структуру: киназному домену предшествует N-концевой домен, а после киназного домена расположен небольшой С-концевой участок. Уровень Aurora А увеличивается на ранней стадии митоза, а при наступлении анафазы он понижается. Снижение содержания фермента, так же как и в случае циклинов, обусловлено его протеолизом с участием убиквитина. Активность Aurora А также регулируется за счет ее фосфорилирования по сайту, расположенному в Т-петле. Как и для Cdk1, это фосфорилирование играет существенную роль в активации Aurora А, и процесс находится под контролем нескольких, пока неполностью охарактеризованных белков. В регуляции активности киназы Aurora А, и, вероятно, Aurora В также участвует протеинфосфатаза 1 (РР1).
Хотя очевидно, что киназы Aurora необходимы для процессов митоза, предстоит еще многое выяснить об их регуляции и о деталях механизма действия. Также неясно, каким образом активность киназ Aurora связана с активностью других киназ, принимающих участие в митозе. Например, неизвестно, функционируют ли они в митозе параллельно, по отдельности или в сочетании с другими киназами.
К числу важнейших выводов, полученных в результате исследований киназ Aurora, относится признание того, что эти ферменты, вероятно, участвуют в генезе опухолей. Опухоли многих типов содержат повышенный уровень киназ Aurora, и гиперэкспрессия Aurora А может приводить к злокачественной трансформации клеток грызунов. Хотя механизм этого явления неизвестен, гиперэкспрессия фермента, вероятно, вызывает амплификацию центросом и дефекты сегрегации хромосом. Более того, считается, что ген, кодирующий Aurora А, обусловливает предрасположенность к развитию рака.
Обобщая все изложенное выше, отметим, что инициация митоза и прохождение клетки через эту стадию цикла требуют функционирования CDK, Plk, NEK и протеинкиназ семейства Aurora. Поскольку активность этих киназ играет критическую роль в поддержании целостности генома, активность каждой киназы регулируется тщательным образом с тем, чтобы избежать несвоевременной активации и обеспечить своевременную ее инактивацию. Накапливаются данные, свидетельствующие о зависимости активности Plk от присутствия других киназ, особенно CDK.
Киназы семейства Polo обладают N-концевым киназным доменом и С-концевым участком,
содержащим два консервативных домена, называемых Polo-домены (РВ1 и РВ2).
Эти домены участвуют в узнавании киназой своих внутриклеточных партнеров.