• Посттрансляционная транслокация происходит независимо от рибосом и SRP
• Посттрансляционная транслокация широко распространена у дрожжей и в меньшей степени у высших эукариот
• Посттрансляционные транслоконы по составу отличаются от котрансляционных транслоконов, однако они представляют собой одни и те же каналы
При котрансляционной транслокации адресование и инициация транслокации белка происходят на очень раннем этапе синтеза новообразованной цепи. Это предотвращает формирование в цитозоле нативной структуры белка, не способной к транслокации. Другой путь транслокации включает полный синтез белка в цитозоле, где его молекула не приобретает нативной структуры, и поэтому может быть транслоцирована.
Такая посттрансляционная транслокация особенно характерна для одноклеточных эукариот, но может происходить и у высших. Эта форма транслокации не зависит от SRP и рибосом и отличается от котрансляционной транслокации по способу реализации и по механизму.
Первым свидетельством в пользу существования второго пути адресования и транслокации было наблюдение о том, что многие белки дрожжей способны к транслокации in vitro, даже после того, как сошли с рибосом. В соответствии с этим наблюдением показано: клетки S. cerevisiae, где отсутствует SRP, жизнеспособны, и многие белки сохраняют способность к транслокации в таких клетках.
Сейчас стало очевидным, что в нормальных условиях для некоторых дрожжевых белков используется только один из двух путей транслокации, однако во многих случаях эффективно используются оба.
Посттрансляционная транслокация характеризуется отсутствием цитозольного фактора, узнающего сигнальные последовательности. Использование того или иного пути транслокации, по-видимому, определяется степенью гидрофобности сигнальной последовательности, которая предоставляет способность взаимодействовать с SRP.
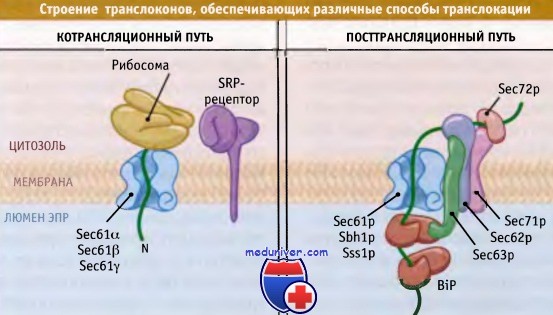
Различные белки взаимодействуют с одним и тем же каналом,
обеспечивая котрансляционную или посттрансляционную транслокацию.
При отсутствии взаимодействия трансляция не задерживается, и белок подвергается адресованию и транслокации уже после окончания синтеза.
Одно из последствий быстрого адресования при котрансляционной транслокации заключается в том, что в цитозоле у субстратов транслокации не образуется нативной структуры. До адресования длина пептида, выходящего из рибосомы, недостаточна для образования нативной структуры, и вместе с тем нативная структура не может образоваться в ограниченном пространстве канала.
Напротив, образование нативной структуры в цитозоле у посттрансляционно транслоцируемых субстратов ингибируется при их взаимодействии с шаперонами семейства hsp70. Эти белки используют энергию гидролиза АТФ для обратимого связывания с посттрансляционно измененными белками, тем самым предотвращая образование их нативной структуры или их агрегацию. При этом белки получают возможность взаимодействовать с каналом.
В посттрансляционной транслокации не участвует такой компонент, как SRP, который узнает сигнальную последовательность до момента ее позиционирования на мембране. Вместо этого белок транспортируется к транслокону с участием мультибелкового комплекса, который является частью посттрансляционного транслокона. Так же как и котрансляционный транслокон, он содержит тримерный комплекс Sec61.
Однако в посттрансляционном транслоконе также присутствуют четыре других белка (Sec62p, Sec63p, Sec71p и Sec72p). Эти белки образуют субкомплекс, большие домены которого открываются в цитозоль и в люмен. Один или несколько этих доменов участвуют в адресовании субстратов, хотя его механизм пока остается невыясненным.
Предполагается, что для транслокации белков через внешнюю мембрану хлоропластов также используется центральный канал, связанный с дополнительными белками. В общем, такой способ переноса белков через мембрану дает возможность гибко регулировать процесс транслокации субстратов и не использовать отдельные каналы для каждого типа белков. Сходным образом регулируется экспрессия генов факторами транскрипции, которые связываются только с промоторами определенных генов, включая и выключая транскрипцию путем взаимодействия с комплексом РНК-полимеразы.
После позиционирования цепи на транслоконе сигнальная последовательность узнается канальными белками, так же как это имеет место при котрансляционной транслокации. Поскольку адресование не зависит от SRP, этот этап приобретает особую важность, так как должен гарантировать отсутствие транслокации белков цитозоля. Так же как и при контрансляционной транслокации, к открытию канала при посттрансляционном переносе приводит узнавание сигнальной последовательности.
По-видимому, способность канала, а не SRP узнавать посттрансляционно позиционированную сигнальную последовательность позволяет предполагать, что, хотя связывание последовательности определяется ее гидро-фобностью, на ее узнавание оказывают влияние другие, пока еще неизвестные факторы.
Хотя система посттрансляционной транслокации широко представлена в клетках S. cerevisiae, данные, свидетельствующие о ее функционировании у высших эукариот, носят менее очевидный характер. Эта форма транслокации пока продемонстрирована in vitro лишь для очень небольших субстратов. В этих случаях образование стабильной нативной структуры в цитозоле маловероятно. Действительно ли эти субстраты in vivo позиционируются посттрансляционно, достоверно не установлено.
Однако у высших эукариот обнаружены гомологи белков Sec62p и Sec63p. Таким образом, возможно, что либо посттрансляционная транслокация происходит несколько по-другому, либо каналы и набор белков используются для других нужд у высших эукариот.
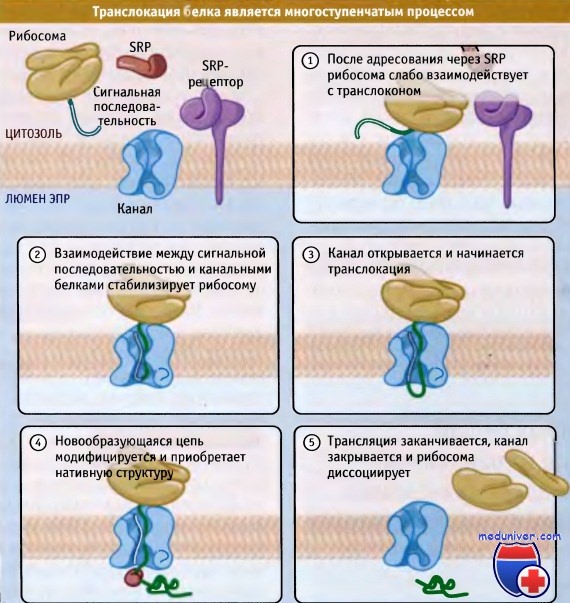
Элонгация новообразующейся цепи в люмен ЭПР не происходит до тех пор,
пока канал не узнал сигнальную последовательность и плотно не связал рибосому.
Вскоре после начала транслокации происходит модификация белковой молекулы,
и она приступает к приобретению нативной структуры.