Перенос глюкозы через мембрану унипортным транспортом
• Для того чтобы преодолеть гематоэнцефалический барьер, глюкоза транспортируется в астроциты через эндотелиальные клетки небольших кровеносных сосудов
• Белки переносят глюкозу за счет унипорта, транспортируя ее в направлении градиента концентрации
• Белки, переносящие глюкозу, претерпевают конформационные изменения, которые приводят к реориентации их субстратных сайтов в мембране клетки
В клетках эукариот основным источником энергии является глюкоза, и многие клетки нуждаются в постоянном ее поступлении, поскольку у них глюкоза служит основным источником энергии для синтеза АТФ. Глюкоза представляет собой полярную молекулу, которая способна к гидратации, а клеточные мембраны не пропускают такие небольшие полярные метаболиты, как сахара. Поэтому для транспорта глюкозы в клетку требуется участие специфических мембранных белков. Транспорт глюкозы через плазматическую мембрану происходит с участием продуктов двух семейств генов.
Переносчики глюкозы (GLUTs) осуществляют независимый перенос (унипорт), при котором через мембрану происходит облегченный транспорт глюкозы. В противоположность GLUT-белкам, котранспортеры Na+/глюкоза расходуют энергию трансмембранного градиента Na+ на транспорт глюкозы. В настоящем разделе мы рассмотрим GLUT-белки.
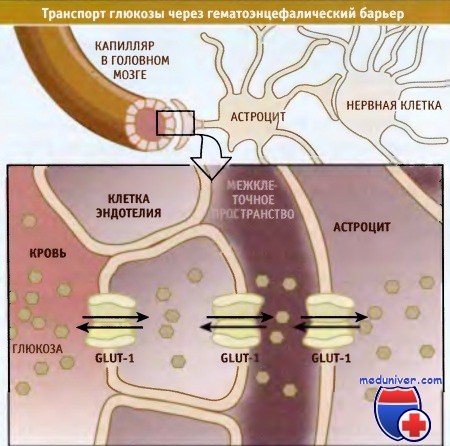
Глюкоза селективно транспортируется через гематоэнцефалический барьер посредством изоформы 1 (GLUT-1) белка-переносчика.
Перенос глюкозы из крови в головной мозг и в другие отделы ЦНС представляет собой многоступенчатый процесс,
в котором участвуют различные типы клеток.
Семейство GLUT-белков является частью более обширного суперсемейства белков, облегчающих основные транспортные процессы (MFS) и широко представленных в клетках всех организмов. GLUT-белки представляют собой интегральные белки мембран клеток эукариот. Изоформы переносчиков GLUT различаются по своим кинетическим характеристикам, специфичностью по отношению к переносимым сахарам, тканевой локализацией и механизмами регуляции. Некоторые GLUT-белки, наряду с глюкозой, транспортируют и другие метаболиты, например галактозу, воду и анальгетики из группы гликопептидов.
GLUT-белки осуществляют унипорт, при котором вещества проходят через мембрану в направлении градиента концентрации. Таким образом, в зависимости от концентрации, GLUT-белки переносят метаболиты в клетку или в противоположном направлении. Поступление в клетку глюкозы, происходящее при участии GLUT-белков, часто определяет жизнеспособность клеток, которые характеризуются высоким уровнем потребления энергии.
Питательные метаболиты, такие как сахара, через кровеносные сосуды транспортируются в органы. Эндотелиальные клетки, выстилающие стенки небольших сосудов, контролируют процессы обмена питательных веществ. В этих эндотелиальных клетках, особенно расположенных в области гематоэнцефалического барьера, содержится много GLUT-белков. Надлежащее функционирование головного мозга сильно зависит от глюкозы, и его клетки особенно чувствительны к снижению ее содержания.
Транспорт глюкозы в нервные клетки происходит через капилляры мозга в несколько этапов и с участием изоформы GLUT-1. Эта изоформа экспрессируется в мембране клеток эндотелия, находящихся на границе между кровью и межклеточным пространством, а также в плазматической мембране астроцитов, функция которых важна в гематоэнцефалическом барьере. Расположенные в этих местах белки GLUT-1 транспортируют глюкозу из крови в эндотелиальные клетки, и оттуда в астроциты. В них глюкоза превращается в другие источники энергии, которые транспортируются в нейроны.
В различных тканях содержатся разные изоформы GLUT. Например, в клетках мышечной и жировой ткани транспорт глюкозы осуществляется с участием GLUT-4. В процессе приема пищи или после еды в клетки этих тканей под действием инсулина увеличивается поступление глюкозы. При этом происходит регулируемый транспорт изоформы GLUT-4 к клеточной поверхности. Эта изоформа также называется инсулин-зависимый переносчик. Белок GLUT-4 локализован во внутриклеточных везикулах, которые сливаются с плазматической мембраной. Этим обеспечивается доставка переносчика GLUT-4 к плазматической мембране и увеличивается емкость транспортного процесса.
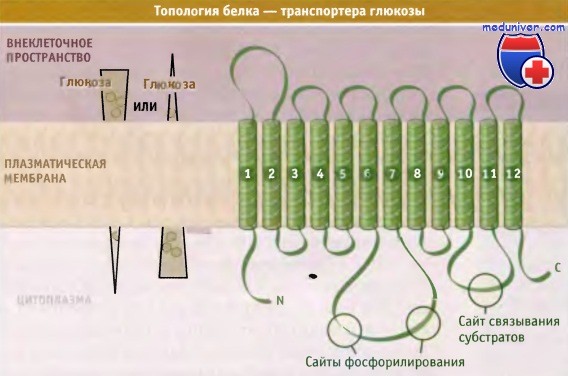
Предполагаемое строение GLUT-переносчика, состоящего из 12 трансмембранных сегментов с внутриклеточными С- и N-концевыми участками.
Внутриклеточные петли содержат сайты фосфорилирования и связывания субстратов.
Показано, что градиент глюкозы по обеим сторонам мембраны может возникать в любом направлении, в зависимости от типа клеток и их метаболизма.
Градиент определяет направление транспорта.
Связывание инсулина со своим поверхностным рецептором запускает каскад внутриклеточных процессов, приводящих к быстрому слиянию этих везикул с мембраной. Это, в свою очередь, приводит к быстрому увеличению транспорта глюкозы в клетку с участием GLUT-4. При диабете типа II не происходит транспорт глюкозы из крови в мышцы и жировую ткань, очевидно, из-за нарушения позиционирования GLUT-4 на плазматической мембране. Более того, GLUT-2 способен экспортировать глюкозу из клеток тех органов, в которых она образуется, например из клеток печени.
По строению переносчик GLUT напоминает другие члены суперсемейства MFS. Предполагается, что он состоит из 12 трансмембранных а-спиралей, имеющих внутриклеточные N- и С-концевые участки и петли. Последние содержат сайт связывания субстрата и сайты фосфорилирования.
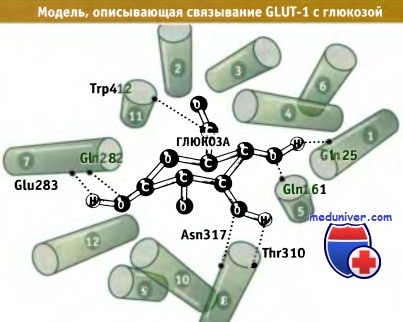
Модель структуры GLUT-1 предложена на основании результатов по сайт-направленному мутагенезу и по измерению транспорта глюкозы мутантным белком. Для построения модели также использовались данные по кристаллической структуре бактериальной лактопермеазы, относящейся к подсемейству переносчиков олигосахари-дов/Н+, входящих в MFS.
Согласно этой модели, при соответствующей ориентации трансмембранных спиралей создается полость поры, через которую транспортируется глюкоза, а также обеспечивается образование водородных связей между GLUT-белком и глюкозой. Результаты кинетического анализа транспорта глюкозы в эритроцитах позволяют предполагать существование специального механизма, который обеспечивает белку GLUT возможность принимать одну из двух основных конформаций. Этот механизм аналогичен постулируемому для бактериальной лактопермеазы. Когда белок находится в одной конформации, сайт связывания глюкозы обращен в сторону внеклеточного пространства. В другой конформации он обращен в сторону цитозоля. При связывании глюкозы с любой стороны происходят конформационные изменения, которые приводят к реориентации сайтов связывания глюкозы к противоположной стороне мембраны и к ее высвобождению.
Таким образом, хотя переносчики GLUT являются унипортерами, а бактериальная лактопермеаза представляет собой симпортер, предполагается, что они функционируют по одному механизму.
Мутации в гене GLUT-1 вызывают значительные дефекты развития. У детей потребность головного мозга в глюкозе в 3-4 раза выше, чем у взрослых, и в мозг поступает до 80% всей глюкозы. Мутации в гене GLUT-1 человека служат причиной проявления редкого синдрома недостаточности GLUT-1, который характеризуется развитием припадков и замедлением развития. Считается, что это связано с нарушением транспорта глюкозы в головной мозг. Эмбрионы мышей, дефектных по гену GLUT-1, отстают в росте, и у них обнаруживаются различные пороки развития. Аналогичные дефекты возникают у мышиных эмбрионов, которые развиваются в организме диабетических самок. При этом избыток глюкозы в крови подавляет экспрессию GLUT-1 в органах эмбриона.
Предполагаемая ориентация трансмембранных а-спиралей переносчика глюкозы GLUT-1.
Показаны остатки, участвующие в связывании глюкозы (размер молекулы глюкозы представлен без учета масштаба).
Вид на спирали со стороны внутренней части плазматической мембраны.
Эта модель построена с использованием гомологичной структуры лактопермеазы E.coli в качестве матрицы.