• Трансмембранные белки должны быть ориентированы с учетом строения мембраны
• Механизм интеграции трансмембранных доменов зависит от типа белка, причем особенно это относится к белкам, которые пронизывают мембрану более одного раза
Процессы узнавания трансмембранных доменов и их интеграция осложняются необходимостью определенной ориентации каждого белка в мембране ЭПР. Для некоторых мембранных белков N-терминальные домены должны располагаться на цитозольной стороне мембраны, а для других требуется противоположная ориентация. Ориентация белка в мембране определяется в ходе процессов его адресования и транслокации.
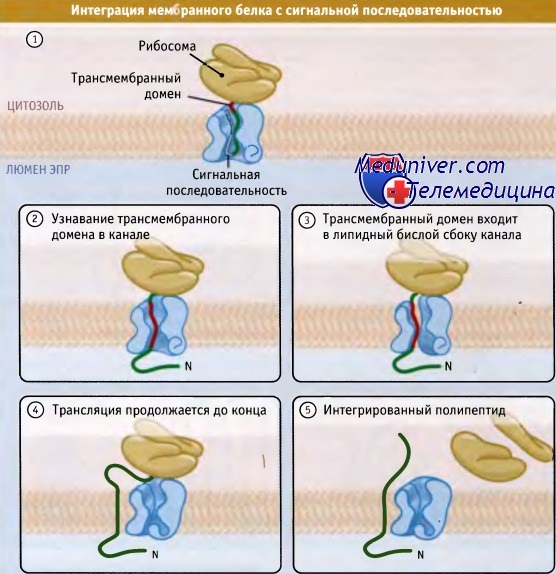
Проще всего понять, каким образом ориентируются белки, которые пронизывают мембрану один раз и в составе которых присутствует отщепляемая N-терминальная сигнальная последовательность. Транслокация этих белков начинается так же, как и секреторных белков, т. е. адресование и инициация транслокации осуществляются с помощью сигнальной последовательности.
Транслокация продолжается до тех пор, пока трансмембранный домен не сойдет с рибосомы, т. е. до начала интеграции. В результате, С-терминальный домен не переносится, а остается с цитозольной стороны мембраны. К числу мембранных белков такого типа относятся рецептор ЛНП и предшественник амилоида.
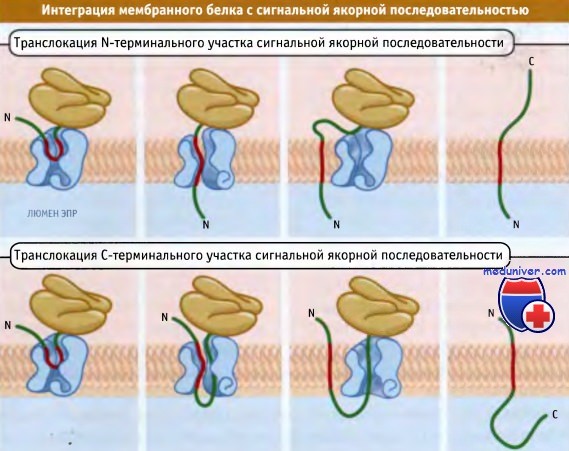
В отличие от трансмембранных белков с отщепляемой сигнальной последовательностью, адресование белков с сигнальной якорной последовательностью происходит с участием внутреннего трансмембранного домена (сигнального якоря). В зависимости от белка трансмембранный домен может находиться в одной из двух ориентаций. От того, как он интегрируется, зависит дальнейшая ориентация всего белка.
Некоторые домены приводят к транслокации С-терминального участка; домены, интегрирующиеся в противоположном направлении, вызывают транслокацию N-терминального участка. Другой класс мембранных белков интегрируется с С-терминального гидрофобного конца, а другая часть белка остается в цитозоле. Поскольку эти трансмембранные домены локализованы на С-терминальных концах белков, они должны узнаваться посттрансляционно.
Сигнальная последовательность инициирует транслокацию,
которая протекает, как и для секреторных белков, до момента трансляции трансмембранного домена и узнавания его в канале.
При этом белки могут интегрироваться только в одной ориентации.
Механизм интеграции этих белков, включая вопрос использования ими транслокационного канала Sec61, исследован недостаточно.
Что определяет ориентацию трансмембранного домена в транслоконе? У бактерий, по-видимому, это определяется распределением заряженных остатков, расположенных по бокам от трансмембранного домена. Заряженные молекулы липидов асимметрично распределяются между двумя слоями плазматической мембраны бактериальной клетки, что объясняет ориентацию. Однако липиды в мембране ЭПР не распределены асимметрично по отношению к заряду, так что это не объясняет ориентацию у эукариот.
Наряду с этим, некоторые белки могут принимать множество топологических форм, что позволяет предположить участие других факторов ориентации. В общем, механизм ориентации трансмембранных доменов у эукариот остается пока неясным.
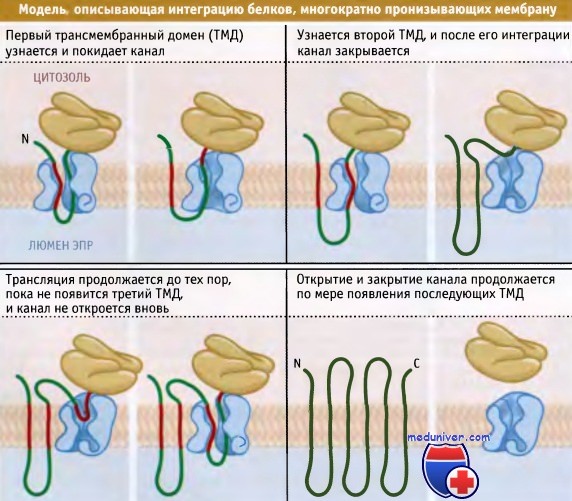
Столь же трудно выявить факторы, управляющие интеграцией и ориентацией политопных белков, т. е. белков, несколько раз пронизывающих мембрану. Простейшая модель предполагает, что трансмембранные домены интегрируются один за другим по мере их появления в канале. Ориентация всего белка определяется свойствами его первого трансмембранного домена. Этот механизм объясняет лишь некоторые случаи, и часто процесс протекает более сложным образом.
Трансмембранные домены не обязательно интегрируются сразу после их узнавания, и в некоторых случаях на транслоконе могут одновременно позиционироваться, по крайней мере, два домена. Эти домены могут интегрироваться парой, а не поодиночке. До интеграции трансмембранные домены в результате взаимодействия друг с другом могут реориентироваться. Таким образом, последовательность событий интеграции политопного мембранного белка носит индивидуальный характер, и его ориентация определяется суммарным влиянием нескольких участков, присутствующих в первичной последовательности.
Интеграция больших белков пока не исследована детально. Однако на основании известных процессов инициации, транслокации и интеграции трансмембранных доменов для небольших мембранных белков предложена соответствующая модель. При исследовании в электронном микроскопе комплекса рибосома-транслокон между рибосомой и каналом обнаружен промежуток величиной около 15°. Узнавание трансмембранного домена в канале, вероятно, тормозит транслокацию, и при этом последующая порция новообразующейся цепи проходит в цитозоль через этот промежуток.
Возможно, что изменение структуры рибосомы облегчает этот процесс. Следующий гидрофобный домен, сходящий с рибосомы, вероятно, имеет высокое сродство к сайту канала, связывающемуся с сигнальной последовательностью/трансмембранным доменом, и цепь опять входит в канал, возобновляя транслокацию. Циклы транслокации приостанавливаются и возобновляются до тех пор, пока не завершится синтез и узнавание всех трансмембранных доменов.
В зависимости от белка, или N- или С-терминальный участок
сигнальной якорной последовательности транслоцируется после ее первоначального взаимодействия с каналом.
Поэтому мембранные белки ориентируются по-разному.
Согласно этой модели, транслокация трансмембранных доменов происходит по очереди.
Ориентация первого домена определяет ориентацию белка в целом.
Показано, что трансляция была инициирована сигнальной якорной последовательностью,
но модель также применима к белкам, позиционированным с помощью сигнальной последовательности, которая после этого отщепляется.