• Локализация митотического веретена определяет место формирования сократимого кольца
• Митотическое веретено удерживается на месте за счет взаимодействия между астральными микротрубочками и кортексом клетки
• Между двумя отдельными группами хромосом в анафазе образуются параллельно расположенные пучки микротрубочек, которые называются структурами межзональной области
• По мере прохождения анафазы, структуры межзональной области сливаются в один большой пучок микротрубочек, называемый остаточным тельцем
• Структуры межзональной области подают кортексу клетки сигнал готовности к образованию сократимого кольца
Взаимодействие между митотическим веретеном и клеточным кортексом определяет место образования сократимого кольца. В ранней анафазе весь кортекс способен поддерживать формирование сократимого кольца и бороздки, однако позже это осуществляется лишь небольшим его участком. Роль веретена в определении выбранной области была продемонстрирована в экспериментах по перемещению веретена на новое место, проведенных на анафазных клетках.
Если в анафазной клетке, которая уже начала формировать сократимое кольцо, переместить веретено на другое место, то в нем начинает формироваться второе сократимое кольцо. Первое кольцо постепенно исчезает, и если веретено не перемещается, клетка будет делиться с участием второго кольца. Такой результат получался независимо от расстояния, на которое удаляли веретено от своего исходного положения, однако если веретено перемещали после прохождения клеткой определенной точки в анафазе, то второе кольцо не образовывалось.
Таким образом, в ранней анафазе весь кортекс способен реагировать на местонахождение веретена, однако позже он утрачивает эту способность.
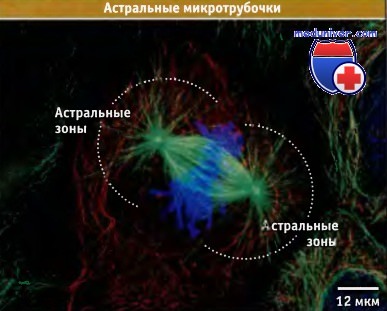
Астральные микротрубочки, отходящие от каждого полюса метафазного веретена.
Микротрубочки окрашены зеленым цветом, хромосомы — синим, а промежуточные филаменты — красным.
Края структурной сети промежуточных филаментов определяют границы клетки.
Отметим, что многие астральные микротрубочки тянутся до плазматической мембраны клетки.
Само веретено удерживается в клетке с помощью астральных микротрубочек. Они распространяются из каждого полюса и достаточно длинные для того, чтобы образовать тесный контакт с большей частью клеточного кортекса.
Как показано на рисунке ниже, в клетках млекопитающих положение веретена зависит от взаимодействия астральных микротрубочек с заякоренным в кортексе динеином. Поскольку динеин движется к минус-концам микротрубочек, локализованным у полюсов веретена, на каждой микротрубочке развивается тянущее усилие. Ориентация веретена определяется локализацией или активацией динеина в кортексе клетки, ее формой, а также относительным количеством астральных микротрубочек, выходящих из полюсов веретена.
Не исключено, что расположение веретена определяется состоянием механического равновесия, которое достигается при уравновешивании тянущих усилий астральных микротрубочек обоих полюсов. Обычно веретено стремится ориентироваться таким образом, чтобы его длинная ось оказалась параллельной длинной оси клетки. Однако существуют исключения.
В клетках некоторых развивающихся тканей цитокинез приводит к образованию двух дочерних клеток, которые сильно различаются по размерам. Такое асимметричное деление является результатом внезапного движения веретена в митозе: в середине анафазы веретено быстро меняет свое расположение и сдвигается ближе к одной из сторон клетки. В результате, бороздка деления смещается в сторону от центра клетки. Что служит причиной такого движения веретена, и почему оно сдвигается в определенное положение, пока остается загадкой.
Каким образом веретено определяет место формирования сократимого кольца? До недавних пор предполагалось, что его локализация определяется астральными структурами. В результате многочисленных экспериментов исследователи пришли к заключению, что, как правило, цитокинез происходит между двумя астральными фигурами, независимо от того, связаны ли они между собой веретеном. Предполагалось, что факторы, необходимые для формирования сократимого кольца, накапливаются на клеточной мембране, в области перекрывания астральных микротрубочек.
В настоящее время, однако, ясно, что положение сократимого кольца не зависит от астральных структур, а определяется другим образованием, состоящим из микротрубочек, которое образуется между индивидуальными группами хромосом в ранней анафазе.
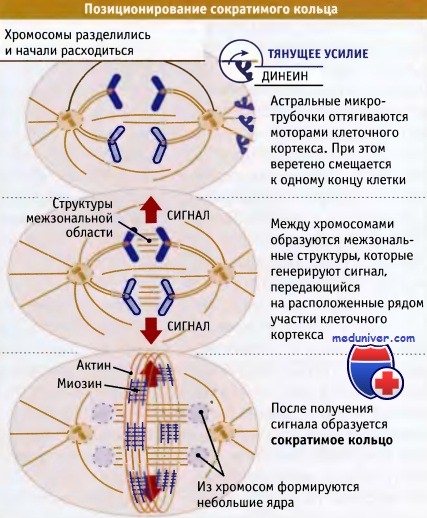
Положение сократимого кольца определяется серией процессов.
Положение веретена в клетке прежде всего зависит от взаимодействия между его астральными микротрубочками и динеином в клеточном кортексе.
Затем структуры межзональной области, которые образуются по мере позиционирования веретена,
продуцируют сигнал, инициирующий сборку сократимого кольца в расположенных рядом участках кортекса.
По мере разделения хромосом, в области, где они выстраиваются в метафазе, образуется много мелких пучков микротрубочек, расположенных параллельно веретену. Эти пучки образуют межзональную область и называются микротрубочками средней зоны (или основными тельцами, stem bodies). Они могут формироваться или из микротрубочек центросом, при их расхождении в анафазе В, или возникать de novo, при полимеризации микротрубочек. Каждый пучок состоит из микротрубочек обеих полярностей, причем плюс-концы микротрубочек противоположной полярности перекрываются в небольшой области в середине пучка. Эти перекрывающиеся области расположены в середине расстояния между полюсами веретена, так, что все пучки центрированы по средней линии.
Перекрывающиеся области имеют специфический состав и содержат уникальный набор белков, включая белковые моторы микротрубочек. В них также локализован кинезинподобный моторный белок, который связывает смежные микротрубочки противоположной полярности (MKLP1, mitosis kinesin-/ike protein 1). Этот белок может принимать участие в образовании структур межзональной области при одновременном движении к плюс-концам двух микротрубочек. Еще один мотор, находящийся в перекрывающейся области, — CENP-E, который до начала анафазы попадает туда из кинетохора.
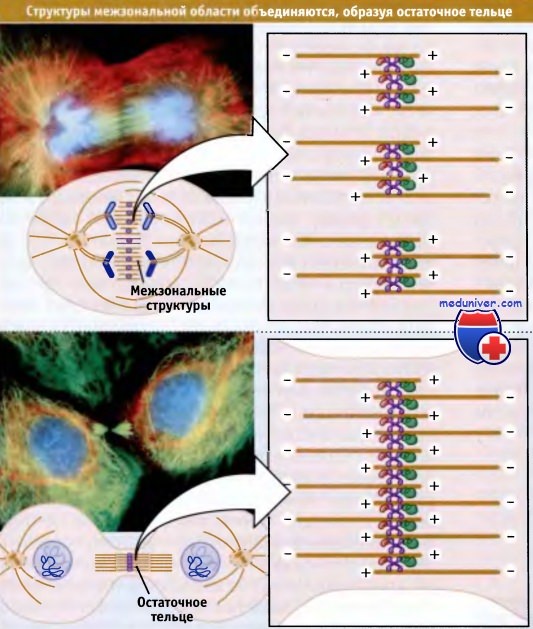
По мере прохождения цитокинеза, отдельные участки межзональной области сливаются с образованием большого крупного пучка микротрубочек, остаточного тельца, которое располагается между двумя изолированными группами хромосом. Также как и у индивидуальных структур межзональной области, центральная область остаточного тельца, в которой перекрываются микротрубочки противоположной полярности, представляет собой специализированную структуру, содержащую большое количество различных белков.
Структуры межзональной области необходимы для формирования сократимого кольца и играют важную роль в определении его местоположения. В сборке сократимого кольца участвует большое количество белков, в том числе его основные структурные компоненты, актин и миозин. Вначале эти белки накапливаются на определенном участке, под плазматической мембраной, где они затем участвуют в сборке кольца. Для накопления и сборки этих белков требуется присутствие структур межзональной области. Если в экспериментальных условиях образование этих структур предотвращалось, то накопления актина и миозина не наблюдалось, и сократимое кольцо не образовывалось.
Также оказалось возможно получить такие клетки, в которых структуры межзональной области находились близко к клеточному кортексу, а не в области митотического веретена. В этих клетках актин и миозин формировали сократимое кольцо в любой точке кортекса, расположенной поблизости от этих структур.
Вероятно, структуры межзональной области также обеспечивают генерацию сигнала, который стимулирует образование сократимого кольца. Молекулярная природа этого сигнала неизвестна, однако возможно, что он возникает в специальной области, находящейся в середине межзональной области, где сосредоточены плюс-концы микротрубочек. Среди белков, присутствующих в этой области, находится белок, активирующий небольшую ГТФазу Rho. При функционировании клетки Rho регулирует образование структур, содержащих актин, и возможно, что постоянное образование в структурах межзональной области активного фермента и его выход оттуда приводят к образованию сократимого кольца из актина и миозина.
На фотографии вверху показана клетка,
находящаяся в стадии между средней и поздной анафазами, ее хромосомы (окрашены синим) только что достигли полюсов.
Между двумя группами хромосом виден широкий пучок структур межзональной области (окрашен зеленым цветом).
Центр каждой структуры не прокрашен, поскольку содержит уплотнение,
состоящее из разных белков, которые препятствуют связыванию красителя с микротрубочками.
На нижней фотографии видны две сестринские клетки, связанные друг с другом только посредством остаточного тельца.
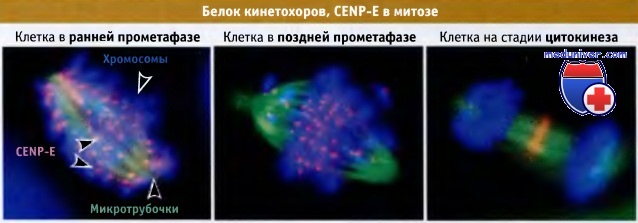
Серия иммунофлуоресцентных микрофотографий, иллюстрирующих, как меняется распределение CENP-E в ходе митоза.
До наступления метафазы и в течение этой стадии CENP-E является компонентом кинетохоров и окрашивается в виде отдельных точек.
Однако в начале анафазы белок выходит из кинетохоров и накапливается в узкой зоне, расположенной поперек экватора веретена, там, где перекрываются микротрубочки.
Когда CENP-E локализуется на ДНК, он окрашивается пурпурным цветом, а когда присутствует на микротрубочках, то оранжевым.