• Центросомы расположены на полюсах веретена и участвуют в его образовании
• Центросомы участвуют в нуклеации микротрубочек и после этого часто остаются связанными с их минус-концами
• Центросомы состоят из двух центриолей, окруженных перицентриолярным материалом
• Для образования новой центросомы необходимо удвоение (дупликация) центриолей
• Дупликация центриолей контролируется клеточным циклом и скоординирована с репликацией ДНК
• Дупликация центриолей происходит в процессе образования и роста новой центриоли, которая с момента образования примыкает к существующей
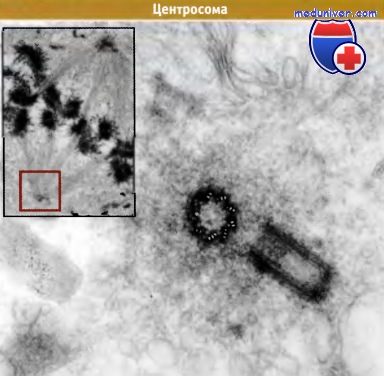
На основной фотографии помещено изображение центросомы в электронном микросокопе.
Две центриоли расположены под прямым углом друг к другу, так, что одна выглядит как круг, а другая как прямоугольник.
Вокруг первой центриоли находится скопление гранулярного материала
(сравните область, примыкающую к центриоли, с более удаленными частями цитоплазмы,
которые окрашены менее интенсивно и где заметно присутствие многих мембранных везикул).
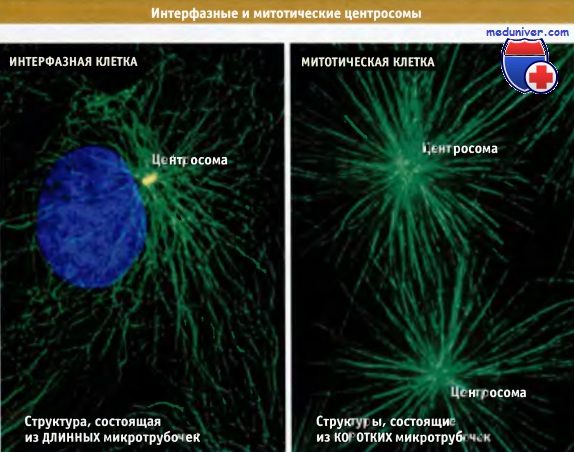
Когда клетка приближается к митозу, в ней происходит ряд изменений. Для клеток млекопитающих наиболее заметные изменения заключаются в исчезновении из цитоплазмы длинных пучков микротрубочек и в появлении двух лучистых астральных структур, состоящих из более коротких микротрубочек, которые часто называют звездами. Этот процесс показан на рисунке ниже. Каждая из радиальных лучистых структур окружает центросому. При расхождении центросом из этих структур в клетках млекопитающих образуется веретено.
Поскольку две центросомы образуют полюса веретена, крайне важно, чтобы в момент митозе в клетке находились только две центросомы. Когда обе астральные структуры начинают взаимодействовать при образовании веретена, его структура стабилизируется хромосомами. Основную роль при этом играют их кинетохоры.
При образовании клетки каждая содержит одну центросому, которая достается ей от одного полюса веретена при предыдущем делении. В неделящихся клетках эта маленькая органелла обычно расположена поблизости от клеточного центра, где она тесно связана с ядром. В интерфазе центросома действует как центр организации микротрубочек. С его участием в цитоплазме создается сеть микротрубочек, которая пронизывает клетку. Эта сеть участвует в организации цитоплазмы и в транспорте различных материалов и органелл внутри клетки.
Центросомы образуют сеть микротрубочек, действуя как сайты их нуклеации. В пределах центросомы микротрубочка начинает расти из кольцеобразного комплекса, содержащего один из типов тубулина, называемый у-тубулином. После того как нуклеация завершилась, минус-конец микротрубочки обычно остается связанным с центросомой. Затем микротрубочка начинает удлиняться или укорачиваться за счет добавления или удаления молекул тубулина к плюс-концу, который расположен на некотором расстоянии от центросомы. В течение разных периодов времени микротрубочки остаются связанными с центросомой и в некоторых клетках отделяются от центросом под действием локализованных в них ферментов.
Связь с центросомой осуществляется посредством нескольких структурных белков, а также моторов, продвигающихся к минус-концу, включая цитоплазматический динеин и белки, относящиеся к группе кинезинов (HSET).
Каждая центросома, находящаяся в клетке, в митозе способна к образованию полюса веретена. Веретено обладает биполярной структурой, поскольку обычно клетки вступают в митоз, обладая двумя центросомами. Если в клетке находится более двух центросом и она вступает в митоз, то будет образовываться веретено, содержащее слишком много полюсов, и потомство таких клеток может характеризоваться анеуплоидностью. Это предотвращается специальными механизмами, которые позволяют центросоме в клеточном цикле реплицироваться только один раз.
Когда эти контрольные механизмы перестают функционировать, образуется слишком много центросом, которые могут привести к образованию генетических дефектов и к развитию раковых клеток и опухолей. Для того чтобы понять, каким образом обеспечивается биполярная структура веретена, необходимо выяснить строение центросомы и механизм контроля ее репликации.
В интерфазных клетках центросома (желтая точка рядом с ядром) нуклеирует образование длинных микротрубочек, которые пронизывают цитоплазму.
В митотических клетках способность центросом к нуклеации микротрубочек возрастает,
и каждая из реплицировавшихся центросом нуклеирует плотные радиальные структуры (звезды),
состоящие из прямых коротких микротрубочек.
Микротрубочки флуоресцируют зеленым цветом, ДНК — синим.
В световом микроскопе в большинстве клеток центросомы выглядят как одна или две точки. Для того чтобы детально исследовать эти органеллы и оценить всю сложность их строения, используют электронную микроскопию (ЭМ). В электронном микроскопе видно, что центросома содержит пару структур, называемых центриоли. Каждая центриоль представляет собой круговую структуру, содержащую девять триплетов микротрубочек, распределенных равномерно по периметру цилиндра диаметром 0,3 мкм.
Еще в 1888 г. Бовери с сотрудниками пришли к выводу, что центриоль, которая была с трудом видна в их световом микроскопе, представляет собой длительно существующую и независимую органеллу, которая образуется только при делении предсуществующей центриоли. Действительно, за редким исключением, новые центриоли образуются только в связи с существующей центриолью. В некоторых клетках тесная связь между двумя центриолями сохраняется на протяжении клеточного цикла. Однако в интерфазе, во многих клетках эта связь нарушается, и две центриоли независимо перемещаются по клетке.
Каждая центриоль связана с областью диффузного материала, которая в электронном микроскопе выглядит непрозрачной. Если смотреть на центриоль со стороны торца, то непрозрачная область расположена вокруг центриоли. Этот перицентриолярный материал состоит из большого количества белков, прикрепленных к каркасу. Как правило, материнская центриоль содержит больше этого материала по сравнению с дочерней, по крайней мере до момента завершения следующего цикла репликации центриоли. Среди белков перицентриолярного материала находится несколько типов белковых моторов микротрубочек и у-тубулиновые кольца, участвующие в нуклеации микротрубочек. Сами центриоли также содержат ряд специфических структур и ферментов, некоторые из которых также присутствуют в перицентриолярном материале.
В интерфазе клеточного цикла центросомы выполняют несколько совместных функций. В большинстве клеток в фазе G1 материнская центриоль инициирует образование длинной тонкой структуры, покрытой мембраной, отходящей от поверхности клетки, которая называется первичная ресничка.
Хотя обычно на них не обращают внимания, первичные реснички можно наблюдать очень часто, и легче перечислить клетки, у которых они отсутствуют, чем те, которые ими обладают. В некоторых эпителиях эти структуры могут простираться более чем на 20 мкм от тела клетки. Поскольку эти структуры не обнаруживаются у многих трансформированных клеток, первичные реснички не играют критической роли в жизни клетки. Этот факт позволил исследователям предположить, что они просто представляют собой рудиментарные образования, подобно аппендиксу.
Однако внешние сегменты палочек и колбочек глаза — высокоспециализированные структуры, в которых происходит поглощение фотонов, образованы производными от первичных ресничек, и накапливаются данные, что первичные реснички также необходимы для процессов нормального развития и функционирования тканей.
В клетках животных количество пар центриолей определяет число центросом. Поэтому клетки контролируют число центросом, которые они содержат, за счет регулирования репликации центриолей. Только начинают появляться работы, в которых вскрываются механизмы контроля за точным удвоением центриолей, и связи этого удвоения с активностью ядра в клеточном цикле (например, с процессом репликации ДНК). В настоящее время очевидно, что наступление репликации центриолей контролируется изменениями в цитоплазме, что указывает на регулирующую роль растворимого фактора. Наряду с этим, условия, позволяющие центри-олям вступить в фазу репликации, существуют только в S-фазе клеточного цикла, когда также реплицируется ДНК.
По-видимому, основным регулятором репликации центросом является CDK2 киназа и ее активаторы циклин А и Е. Эти регуляторы активируются перед началом S-фазы и также контролируют вхождение клетки в фазу синтеза ДНК. То, что один и тот же регулятор контролирует репликацию ДНК и центриолей, обеспечивает скоординированность двух этих процессов, так что клетка входит в митоз по завешении репликации центросомы и хромосом. Хотя неизвестно, как инициируется дупликация центриолей, не вполне ясно, каким образом она ограничивается образованием одной новой центриоли из каждой предсуществующей.
Начавшись, дупликация центриолей приводит к постепенному образованию новой центриоли, примыкающей к каждой из двух, с которыми клетка вступила в S-фазу. Одна из них называется дочерней и является более молодой, поскольку образовалась в предыдущем клеточном цикле. Другая, материнская, образовалась в раннем цикле и характеризуется наличием первичной реснички. Первый признак репликации центриоли состоит в появлении двух коротких процентриолей, каждая из которых распространяется под прямым углом к стенке одной из центриолей.
Этот процесс не зависит от физического взаимодействия между двумя исходными центриолями, поскольку происходит, даже когда они разобщены. После образования процентриолей они медленно элонгируют до тех пор, пока их длина не достигнет размеров зрелой центриоли к моменту митоза. Хотя в случае материнской и дочерней центриоли процентриоли образуются и растут одинаковым образом, большая часть перицентриолярного материала остается связанным с более зрелой, материнской центриолью.
В конце концов, в процессе дупликации дочерняя центриоль накапливает новый перицентриолярный материал; частично это происходит за счет микротрубочек, которые она организует. В поздней интерфазе клетка содержит две центросомы, каждая из которых состоит из пары тесно связанных друг с другом центриолей и перицентриолярного материала. В некоторых клетках две центросомы остаются физически связанными между собой и функционируют как единое целое до момента вступления в митоз. В других клетках связь между центросомами разрушается, и они расходятся до момента вступления клетки в митоз. Время наступления момента разделения двух центросом по отношению к разрыву ядерной оболочки сильно варьирует, даже среди генетически однородных клеток, растущих в тесном контакте.
Митоз в клетке кенгуровой крысы. На левом снимке видны четыре центросомы (помечены желтыми стрелками).
Образуются три метафазных пластинки (желтые линии на центральном снимке)
и четыре группы хромосом (помечены стрелками на правом снимке).
При цитокинезе образуются четыре анеуплоидных клетки.
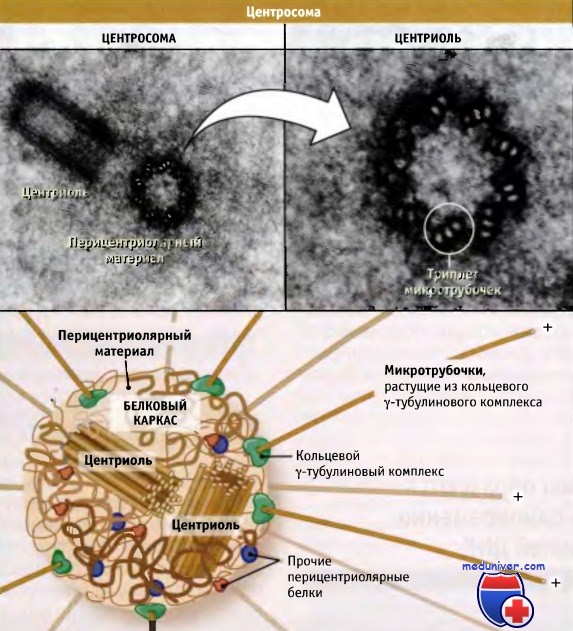
В митотической клетке (вверху слева) центросома состоит из двух центриолей
— материнской (на этой микрофотографии представлена в поперечном сечении) и дочерней (представлена в продольной проекции)
— окруженных темной областью аморфного перицентриолярного материала.
Каждая центриоль (наверху справа) состоит из триплетов микротрубочек, образующих как бы стенку цилиндра.
На рисунке внизу видно, что центриоли расположены под прямым углом по отношению друг к другу.
Вокруг них находится много различных белков, образующих перицентриолярный материал.
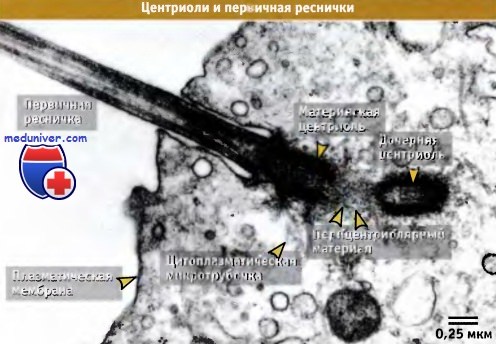
Электронная микрофотография основания первичной реснички.
Структура растет от материнской центриоли, которая находится под плазматической мембраной.
Между материнской и дочерней центриолями находится перицентриолярный материал плотного гранулярного вида.
Показана лишь часть клетки и первичной реснички.
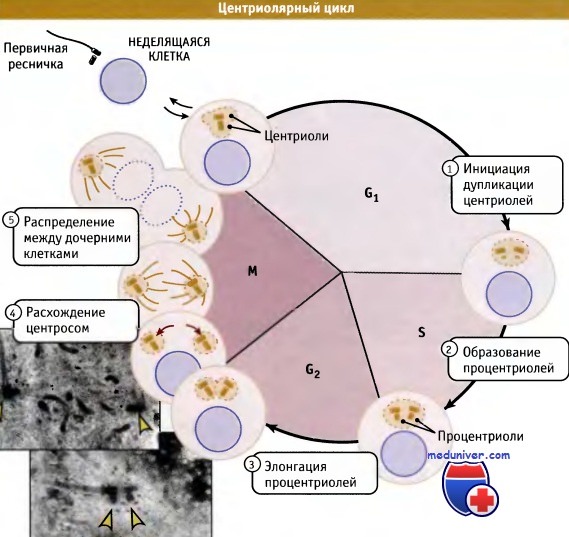
Центриолярный цикл в клетках млекопитающих.
Две центриоли центросомы материнской клетки дуплицируются, и затем две образующихся центросомы распределяются по дочерним клеткам.
На вставке представлены две микрофотографии реплицированных центросом до и после их расхождения в начале митоза.