Как было отмечено ранее, клеточные оболочки грамотрицательных и грамположительных бактерий представляют собой различные структуры. Основное различие в составе их клеточных стенок состоит в том, что по сравнению с грамотрицательными бактериями грамположительные характеризуются более толстым слоем пептидогликана. Кроме того, наряду с пептидогликаном, в клеточной стенке грамположительных бактерий содержится еще один важный класс макромолекулярных полимеров.
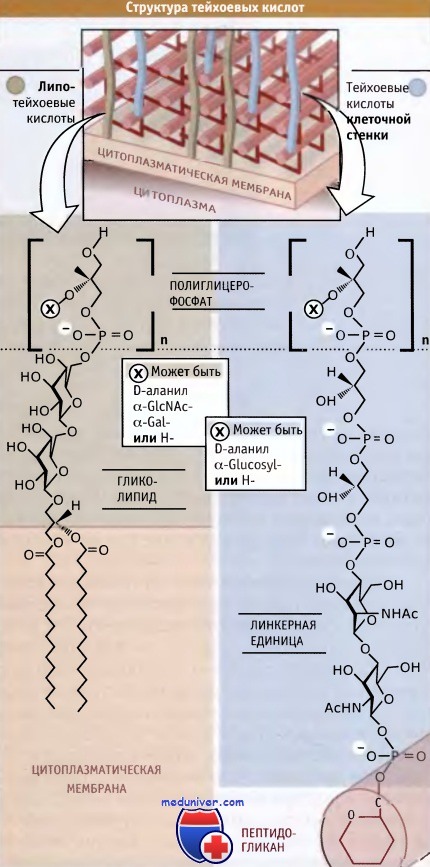
Как правило, эти полимеры являются полианионами. К числу хорошо известных и наиболее изученных из них принадлежат тейхоевые кислоты. Рисунок ниже иллюстрирует, что тейхоевые кислоты имеют простую повторяющуюся структуру, состоящую из полимера, содержащего фосфорилированные формы сахаров или остатков глицерина. Связывание полимера с клеткой происходит одним из двух способов. Липотейхоевые кислоты связываются с остатками жирных кислот цитоплазматической мембраны, а тейхоевые кислоты клеточной стенки связываются с пептидогликаном посредством линкерных групп.
В разных бактериях и даже в родственных штаммах одного вида обнаружено огромное количество различных повторяющихся единиц. Полимеры имеют общее свойство — все они являются полианионами, что обусловливается наличием в их составе фосфатных групп. Однако фосфат не играет критической роли, и его отсутствие может заменяться другой группой, например глюкоуронатом. При этом образуется тейхоуроновая кислота. Так же как и пептидогликаны, тейхоевые кислоты синтезируются в цитоплазме, через предшественник, связанный с УДФ, и затем транспортируются вне клетки для сборки.
Тейхоевые кислоты необходимы для жизнедеятельности клетки. Об этом свидетельствует тот факт, что делеция генов, участвующих в их синтезе, приводит к гибели бактерий. Полностью их функции не выяснены, хотя не исключено, что они каким-то образом связаны с поддержанием на клеточной стенке электрического заряда. Также предполагается, что они участвуют в захвате клеткой двухвалентных катионов, контролируют активность ферментов автолиза, действующих на пептидогликан, или обеспечивают общую проницаемость клеточной стенки.
Эксперимент, демонстрирующий постепенное построение новой клеточной стенки у В. subtilis при переходе от среды, дефицитной по магнию, к среде с недостатком фосфата.
Предсуществующая клеточная стенка (содержащая тейхоевую кислоту) постепенно,
на всем протяжении равномерно замещается новосинтезированным материалом (содержащим теихоуроновую кислоту).
У грамположительных бактерий с отсутствующей наружной мембраной существует два механизма удержания поверхностных белков, с тем чтобы не допустить их потери. Один механизм представлен на рисунке ниже.
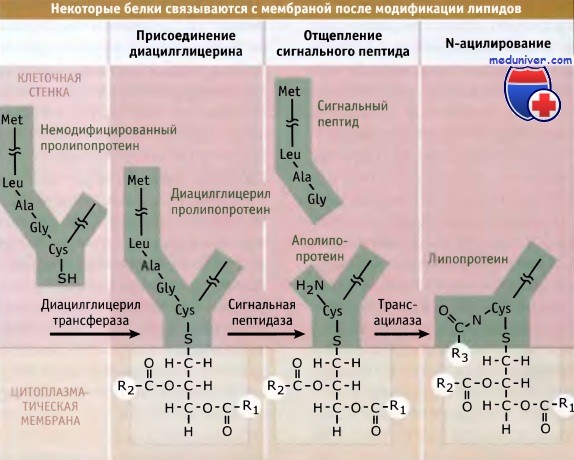
Набор белков, секретируемых по общему секреторному пути (Sec), подвергается расщеплению специфической сигнальной пептидазой «типа II». До их расщепления фермент фосфатидилг-лицерин: пролипопротеин диацилглицерил трансфераза присоединяет белок к фосфофипидам на наружной стороне цитоплазматической мембраны. Белки, предназначенные для этой «липомодификации», узнаются по наличию сигнала сортинга, который содержит цистеиновый остаток сразу же после сайта расщепления сигнальной пептидазой. Ковалентное связывание белка с наружной поверхностью цитоплазматической мембраны предотвращает его отщепление.
У грамположительных бактерий большое количество белков подвергается модификации с участием такого механизма. Например, считается, что у В. subtilis существует более 100 подобных белков. (Хотя у грамположительных бактерий этот механизм чаще используется для поверхностно-связанных белков, липомодифицированные белки также присутствуют у грамотрицательных бактерий.)
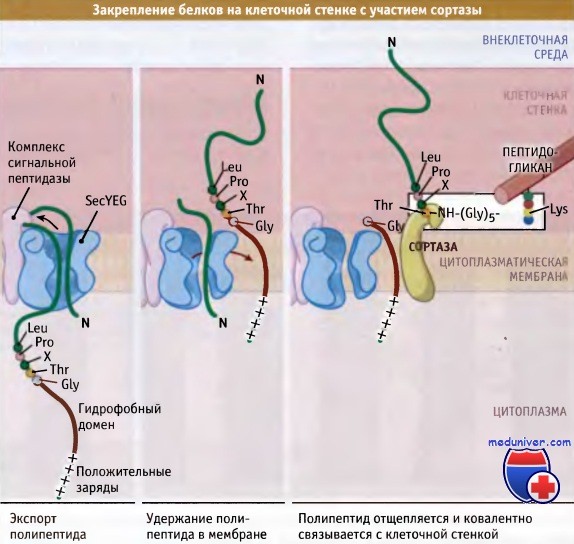
Второй механизм удержания белков на клеточной поверхности у грамположительных бактерий состоит в связывании белка с пептидогликаном клеточной стенки. Вообще говоря, многие белки, связанные с клеточной стенкой, по-видимому, участвуют во взаимоотношениях между патогеном и организмом хозяина (хотя это предположение может являться следствием систематической ошибки секвенирования генома патогенного микроорганизма). Ключевым ферментом, принимающим участие в этом процессе, является сортаза. Известно большое количество субстратов этого фермента.
Наиболее хорошо изучен белок А из Staphylococcus aureus. У белков, прикрепленных к клеточной стенке, на N- и С-концевых участках находятся узнаваемые последовательности. На N-концевом участке присутствует классический расщепляемый сигнальный пептид, необходимый для экспорта белка через цитоплазматическую мембрану. На С-конце, за вторым гидрофобным доменом расположены несколько в основном положительно заряженных аминокислотных остатков. По-видимому, во время работы сортазы этот домен поддерживает С-концевой участок в непосредственной близости от цитоплазматической мембраны.
Сайт узнавания сортазой расположен непосредственно перед С-концевым гидрофобным участком и представляет собой короткую последовательность, обычно Leu-Pro-X-Thr-Gly, где X может быть любая аминокислота. В белке расщепляется связь между четвертым и пятым остатками этой последовательности аминокислот, и вероятно, при действии сортазы, он связывается с пептидогликаном амидной связью, хотя пока такой механизм однозначно не продемонстрирован.
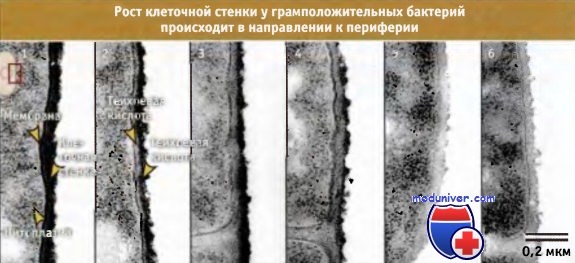
Как иллюстрирует рисунок ниже, синтез клеточной стенки у грамположительных бактерий сопровождается наращиванием нового материала, которое происходит в направлении изнутри-кнаружи. По мере созревания пептидогликана, он мигрирует к внешней стороне, пока не достигнет поверхности, где высвобождается за счет автолиза. Согласно современным представлениям, пептидогликан в релаксированном состоянии включается в состав стенки и, по мере миграции кнаружи, за счет роста клетки, постепенно растягивается. В конце концов, по мере увеличения растяжения, материал оказывается под нагрузкой, и в этот момент наступает гидролиз, после чего продолжается дальнейшее построение клеточной стенки.
Остается неразрешенным важный вопрос, каким образом ферменты автолиза регулируют созревание пептидогликана и рост клетки? Также не ясно, насколько модель синтеза пептидогликана «3 вместо 1», объясняющая наращивание клеточной стенки у грамотрицательных бактерий, приложима к многослойной клеточной стенке грамположительных бактерий.
Даже у таких хорошо известных микроорганизмов, как B.subtilis и E.coli, детали строения клеточной стенки исследованы недостаточно. Причем это несмотря на то, что в основе действия наиболее распространенных антибиотиков (особенно р-лактамов и гликопептидов) лежит ингибирование синтеза клеточной стенки бактерий. В основном это связано с тем, что внимание исследователей было сосредоточено на изучении биохимии взаимодействия ингибиторов с препаратами очищенных ферментов. К числу неизученных относится важнейший вопрос, касающийся механизма синтеза пептидо-гликана с участием РВР, учитывая трехмерную структуру клеточной стенки растущей бактерии.
Для разных микроорганизмов структура клеточной стенки существенно варьирует. Два крайних случая подобной специализации можно проиллюстрировать микоплазмой, у которой вообще отсутствует клеточная стенка, и микобактериями, для которых характерно присутствие сложных жирных кислот, ковалентно связанных с пептидогликаном.
Тейхоевые кислоты являются компонентами клеточной стенки грамположительных бактерий.
Они закрепляются на поверхности клетки либо через цитоплазматическую мембрану, либо через пептидогликан.
У грамположительных бактерий один из механизмов привязки белков
с клеточной поверхностью заключается в присоединении их к фосфолипидам плазматической мембраны.
Жирные длинноцепочечные кислоты обозначены буквами R1, R2 и R3.
У грамположительных бактерий некоторые белки клеточной поверхности связываются с пептидогликаном клеточной стенки.
Предполагается, что фермент сортаза расщепляет связь Thr-GLy и ковалентно закрепляет белок на клеточной стенке.