Строение и функционирование натрий-калиевой АТФазы
• Na+/K+-АТФаза относится к АТФазам P-типа, близким к Са2+-АТФазе и Н+-АТФазе
• Na+/K+-АТФаза поддерживает градиент Na+ и К* через плазматическую мембрану
• Na+/K+-АТФаза плазматической мембраны является генератором электрического заряда: она транспортирует из клетки три иона Na+ на каждые два иона К+, которые она закачивает в клетку
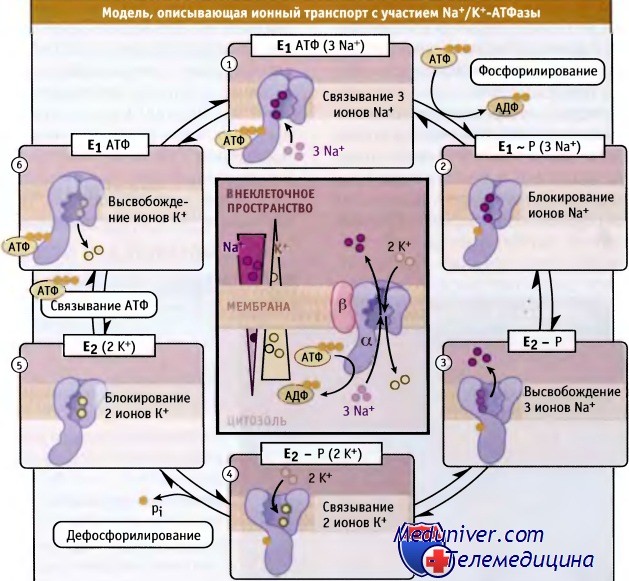
• Рабочий цикл Na+/K+-АТФазы описывается схемой Поста-Альберса, согласно которой фермент совершает оборот между двумя основными конформациями
По отношению к окружающей среде все клетки заряжены отрицательно. Это обусловлено присутствием небольшого избытка положительно заряженных молекул во внеклеточном пространстве и противоположной ситуацией в цитозоле. Для нормального функционирования клетки по сторонам плазматической мембраны необходимо наличие электрохимического градиента.
В этом отношении клетка напоминает электрическую батарею с разделенными зарядами, которую можно использовать для выполнения работы. В клетках млекопитающих градиенты концентрации Na+ и К+ представляют собой два основных компонента трансмембранного электрохимического градиента. Внутри клетки, по сравнению с внеклеточной средой, поддерживается более низкая концентрация ионов Na+ и более высокая ионов К+.
Образование и поддержание электрохимического градиента ионов Na+ и К+ в клетках животных происходят с участием Na+/K+-АТФазы, которая представляет собой ионный насос, использующий энергию гидролиза АТФ для транспорта катионов. С помощью этого фермента в клетке устанавливается отрицательный мембранный потенциал покоя, с помощью которого контролируется необходимый уровень осмотического давления, который не позволяет клетке лизироваться или сжиматься и который также обеспечивает Na+-зависимый вторичный транспорт молекул.
Na+/K+-АТФаза относится к группе АТФаз Р-типа, включающей также Са2+-АТФазу саркоплазматического ретикулума, которая рассматривалась в отдельной статье на сайте (рекомендуем пользоваться формой поиска на главной странице сайта).
АТФазы P-типа представляют собой ферменты, которые при аутофосфорилировании остатка аспарагиновой кислоты в процессе ионного транспорта образуют фосфорилированный промежуточный продукт. В процессе аутофосфорилирования АТФаз P-типа у-фосфатная группа АТФ переносится на активный центр фермента. На каждую гидролизованную молекулу АТФ обмениваются три иона Na+ из цитозоля и два иона К+ из внеклеточной среды. Na+/K+-АТФаза функционирует со скоростью 100 оборотов в 1 с.
По сравнению с потоком ионов через поры каналов такая скорость транспорта представляется низкой. Транспорт через каналы происходит со скоростью 107-108 ионов в 1 с, т. е. близкой к скорости диффузии ионов в воде.
Схема Поста-Альберса для рабочего цикла Na+/K+-ATФаза.
Макроэргическая фосфатная связь обозначена как Е1-Р.
На рисунке в центре представлен весь цикл работы фермента.
Показаны градиенты ионов Na+ и К+ по сторонам плазматической мембраны покоящейся животной клетки.
Были охарактеризованы основные этапы цикла ферментативного транспорта ионов, происходящего с участием Na+/K+АТФазы. Они представлены на схеме Поста-Альберса. Первоначально эта схема была предложена для Na+/K+-АТФазы, а затем использовалась для идентификации специфических состояний молекулы всех АТФаз P-типа. Согласно схеме Поста-Альберса, АТФазы P-типа могут принимать две различные конформации, которые обозначаются как фермент 1 (Е1) и фермент 2 (Е2). Находясь в этих конформациях, они способны связывать, захватывать и транспортировать ионы. Эти конформационные изменения происходят за счет реакции фосфорилирования-дефосфорилирования:
• В конформации внутриклеточный АТФ и ионы Na+ с высоким сродством связываются с АТФазой. При этом фермент переходит в состояние Е1АТФ(3Nа+), происходит АТФ-зависимое фосфорилирование остатка аспарагиновой кислоты и захват трех ионов Na+ в конформации E1 - P(3Na+).
• Дальнейшее изменение конформации приводит к образованию состояния Е2-Р, снижению сродства к ионам натрия, и к выходу их во внеклеточное пространство. Сродство фермента к ионам К+ повышается.
• Связывание с АТФазой ионов К+, находящихся во внеклеточном пространстве, приводит к дефосфорилированию Е2-Р(2К+) и к захвату двух ионов К+ с переходом в состояние Е2(2К+).
• При связывании внутриклеточного АТФ конформация изменяется и ионы К+ отщепляются. При этом возникает состояние Е1АТФ, а связывание внутриклеточного натрия приводит к конформации Е1АТФ(3Na+).
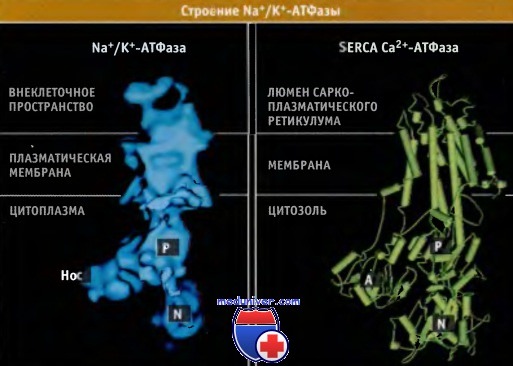
Анализ первичной структуры белков позволяет высказать предположение, что все АТФазы P-типа обладают одинаковой пространственной структурой и транспортным механизмом. Na+/K+-АТФаза состоит из двух субъединиц, каталитической а, одинаковой для всех АТФаз P-типа, и регуляторной субъединицы, b, которая специфична для каждой АТФазы. Более мелкая субъединица b имеет один трансмембранный домен, который стабилизирует а-субъединицу и определяет ориентацию АТФазы в мембране. В клетках некоторых тканей активность Na+/K+-АТФазы, вероятно, регулируется еще одним белком, субъединицей у. Каталитическая субъединица а содержит сайты связывания для АТФ, а также для ионов Na+ и К+.
Эта субъединица в изолированном виде способна осуществлять транспорт ионов, как показано в экспериментах по гетерологичной экспрессии и в электрофизиологических исследованиях.
Структура а субъединицы Na+/K+-АТФазы, построенная по данным криоэлектронной микроскопии, напоминает строение Са2+-АТФазы SERCA. Подобно насосу SERCA, эта субъединица состоит из 10 трансмембранных а спиралей. Внутриклеточный P-домен, расположенный между трансмембранными сегментами 4 и 5, содержит сайт фосфорилирования, который обладает общей структурой для всех АТФаз P-типа. Этот сайт представлен остатком Asp376 в характерной последовательности Asp-Lys-Thr-Gly-Thr-Leu-Thr. Связывание АТФ и ионов Na+ индуцирует существенные изменения конформации петли, соединяющей N- и Р-домены. Эти изменения приводят к сближению сайта связывания АТФ на N-домене и сайта фосфорилирования на Р-домене.
Na+/K+-АТФаза является ионным насосом-генератором. В обычных физиологических условиях свободная энергия гидролиза АТФ (ΔGАТФ) расходуется на транспорт из клетки трех ионов Na+ в обмен на два иона калия, причем ионы переносятся против градиента их концентраций. Таким образом, клетка теряет суммарный положительный заряд. Это способствует росту отрицательного заряда цитозоля по сравнению с внеклеточной средой. В результате по сторонам клеточной мембраны возникает разность потенциалов и осмотический ионный градиент.
АТФазы P-типа представляют собой ионные насосы, использующие энергию гидролиза АТФ для поддержания трансмембранного ионного градиента. Поскольку каждый этап ферментативного цикла носит обратимый характер, АТФазы P-типа в принципе могут продуцировать АТФ, используя энергию трансмембранного потенциала. Таким образом, Na+/К+-АТФаза обладает определенной возможностью функционировать в противоположном направлении. При этом ионы Na+ будут поступать в клетку, а ионы К+ выходить оттуда, что приведет к тому, что поток ионов будет преимущественно направлен в клетку.
Обычный транспорт ионов Na+ из клетки и ионов К+ в клетку происходит, пока величина ΔGАТФ превышает электрохимическую энергию соответствующего ионного градиента. Когда энергия, необходимая для активного транспорта ионов Na+ и К+, становится равной ΔGАТФ, поток ионов прекращается. Эта величина представляет собой потенциал обращения функционирования Na+/К+-АТФазы, т. е. значение мембранного потенциала, ниже которого фермент начинает работать в обратном направлении. Значение потенциала обращения составляет порядка -180 мВ, т. е. он представляет собой гораздо более отрицательную величину, чем мембранный потенциал любой клетки в физиологических условиях. Поэтому маловероятно, что в клетку может поступать поток ионов Na+, имеющий для нее опасные последствия.
Однако все может измениться при снижении кровоснабжения, например при инфаркте миокарда или при интоксикациях, приводящих к недостатку АТФ или к увеличению крутизны ионных градиентов. В конце концов это может вызвать перемену направления транспорта ионов Na+/K+-АТФазой и гибель клетки.
Na+/K+-АТФаза является мишенью для многих токсинов и лекарственных препаратов. Например, растительные стероиды, называемые сердечными гликозидами, такие как уабаин и дигиталис, являются специфическими ингибиторами ионного транспорта, осуществляемого Na+/K+-АТФазой. К числу специфических ингибиторов относятся также другие токсины, например палитоксин из некоторых морских кораллов и сангвинарин из растений. В отличие от сердечных гликозидов, которые подавляют поток ионов через Na+/K+-АТФазу, палитоксин и сангвинарин блокируют АТФазу в открытой конфигурации.
Тем самым ионы получают возможность транспортироваться в направлении их концентрационных градиентов, что приводит к нарушению электрохимических градиентов. Сердечные гликозиды обратимо связываются с сайтами Na+/K+-АТФазы, расположенными вне клетки, при этом ингибируется гидролиз АТФ и транспорт ионов. Тщательно контролируемое ингибирование Na+/K+-АТФазы клеток миокарда сердечными гликозидами, например дигиталисом, применяется при лечении сердечной недостаточности. Частичное ингибирование субпопуляции Na+/K+-АТФаз сердечными гликозидами несколько увеличивает внутриклеточную концентрацию ионов Na+, что приводит к повышению концентрации ионов Са2+, благодаря транспорту через Na+/Са2+-антипортер. Известно, что небольшое увеличение внутриклеточной концентрации ионов кальция усиливает сократимость сердечной мышцы.
Строение а-субъединицы Na+/К+-АТФазы напоминает Са2+-АТФазу саркоплазматического ретикулума (SERCA).
Структура субъединицы установлена на основании данных крио-электронномикроскопического исследования.
Изображение кристаллической структуры SERCA в конформации Е2 получено на основании данных Protein Data Bank file 1IW0.