Модификации промежуточных филаментов после трансляции
• Промежуточные филаменты представляют собой динамичные образования и способны к быстрым периодическим изменениям структуры
• Белки головных и хвостовых доменов подвергаются нескольким посттрансляционным модификациям
• Основным механизмом, обеспечивающим в клетке изменения структуры промежуточных филаментов, является фосфорилирование белков
• Протеолитическая деградация приводит к изменению количеств белков и способствует апоптозу
Несмотря на существование центра организации микротрубочек, мы не располагаем данными о наличии в клетке аналогичного центра для промежуточных филаментов, который инициирует процесс их сборки. In vitro белки промежуточных филаментов собираются легко, без участия кофакторов, и неполярная природа филаментов свидетельствует о том, что сборка может происходить с любого конца.
В клетках свободные концы длинных промежуточных филаментов наблюдаются чрезвычайно редко. Однако фрагменты их можно наблюдать, используя флуоресцентные зонды. Особенно много таких фрагментов находится на периферии клетки, что позволяет предполагать, что в основном там происходит сборка или разборка промежуточных филаментов.
Белки промежуточных филаментов модифицируются посредством фосфорилирования, гликозилирования, фарнезилирования и трансглутаминирования головных и хвостовых доменов, а также подвергаются ферментативной деградации. Однако влияние большинства этих модификаций на функции промежуточных филаментов исследовано мало.
Изменения структуры промежуточных филаментов, связанные с необходимостью реформирования их сети или перераспределения белков в клетке, требуют локальной контролируемой разборки и повторной сборки. Одним из событий, во время которого это происходит, является клеточное деление. Фосфорилирование белков промежуточных филаментов играет важную роль в митозе.
Фосфорилирование белков ламины (типов А- и В-) митотической киназой Cdk1 запускает процесс деградации ядерной мембраны. Вместе с тем, в митозе, промежуточные филаменты цитоплазмы должны на какое-то время приобретать более рыхлую структуру, с тем чтобы обеспечить разделение клетки на две. Белки типа I, II и III фосфорилируются в профазе и при цитокинезе; виментин, GFAP, нестин и К18 содержат общий сайт фосфорилирования киназой Cdk1. Другие киназы также могут участвовать в фосфорилировании.
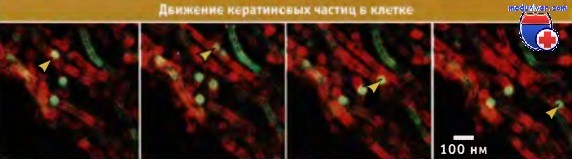
Движение белков промежуточных филаментов по различным элементам цитоскелета, в цитоплазме, вероятно, происходит с помощью моторных белков.
В процессе съемки кератиновая частица, меченная зеленым флуоресцирующим белком (указана стрелкой),
перемещается в поле зрения по микротрубочке (красного цвета), которая частично находится в плоскости поля зрения.
В культуре некоторых эпителиальных клеток можно отчетливо наблюдать деградацию цитоплазматических филаментов в момент вступления клетки в митоз. Она проявляется в виде перераспределения кератинов из филаментов в агрегаты, которые затем, после цитокинеза, снова образуют филаменты. В тканях промежуточные филаменты цитоплазмы неполностью деградируют с образованием агрегатов; они лишь становятся более рыхлыми или деградируют локально, вдоль борозды деления.
Фосфорилирование также является основным механизмом, посредством которого осуществляются изменения структуры промежуточных филаментов в отстутствие клеточного деления. Регуляция сборки и разборки промежуточных филаментов зависит не от гидролиза нуклеотидов (как это имеет место для актина и тубулина), а от процессов фосфорилирования и дефосфорилирования. Во всех группах белков промежуточных филаментов идентифицированы многочисленные сайты фосфорилирования, содержащие серин, и для некоторых белков известны соответствующие киназы.
Однако полностью природа субстратов и ферментов установлена лишь для некоторых белков промежуточных филаментов. В основном сайты фосфорилирования расположены в деспирализованных участках головных и хвостовых доменов, где любое увеличение заряда благоприятствует разборке филаментов. Так, фосфорилирование приводит к росту отрицательного заряда и снижает интенсивность сборки филаментов. Простые кератины фосфорилируются в ответ на возникновение стрессорных условий и с участием активирующихся при этом киназ.
Нестин фосфорилируется при дифференцировке миобластов, что приводит к изменению его экспрессии. Хвостовые домены нейрофиламентов фосфорилируются очень интенсивно; считается, что это является определяющим для создания архитектуры аксона, поскольку отрицательный заряд, возникающий на хвостовых доменах, приводит к их взаимному отталкиванию под прямым углом к оси филамента. За счет фосфорилирования также регулируется взаимодействие белков промежуточных филаментов с некоторыми сигнальными молекулами.
Прочие посттрансляционные модификации, свойственные белкам промежуточных филаментов, включают фарнезилирование и миристоилирование ламинов для их заякоривания на мембране, трансглутаминирование некоторых кератинов, образование дисульфидных связей и гликозилирование. Все эти модификации большей частью носят тканеспецифический характер. Трансглутаминирование кератинов происходит в эпидермальных кератиноцитах и в волосяных фолликулах. В клетках эпидермиса эта модификация определяет ороговение оболочки терминально дифференцированных кератиноцитов, благодаря которой они приобретают устойчивость в физическим воздействиям. Трансглутаминирование также способствует образованию жестких клеточных структур в концентрических слоях формирующегося волоса.
Присутствие цистеина нехарактерно для белков промежуточных филаментов, однако у некоторых кератинов, при созревании жестких кератинсодержащих структур, и при терминальной дифференцировке кератиноцитов, наблюдается образование дисульфидных связей. В белках К18, К13, NF-M и NF-L обнаружены остатки N-ацетилглюкозамина, что свидетельствует об их гликозилировании. У белка К18 этот процесс, вероятно, связан с его растворимой или деполимеризованной формой и позволяет сформировать депо мономеров.
Как следует из экспериментов по трансфекции, протеолиз промежуточных филаментов играет важную роль в удалении апоптотических клеток и помогает регулировать количественный баланс между двумя кератинами, экспрессирующимися совместно. Кератиновые филаменты могут образоваться только из гетеродимеров типа I—II, и если в клетке образуется избыток одного из типов кератина, то белок удаляется путем протеолиза. Белки промежуточных филаментов убиквитинируются для последующего протеолиза, и накопление таких белков характерно для самых различных патологий, от нейродегенеративных заболеваний до цирроза печени. Так же как и фосфорилирование, убиквитинилирование происходит при воздействиях, вызывающих клеточный стресс.
Белки промежуточных филаментов являются мишенями каспаз в процессе клеточной гибели по механизму апоптоза. Сайты чувствительности к каспазе обнаружены в виментине, ламинах и К18. Судьба промежуточных филаментов при апоптозе подробнее всего исследована для ламинов и кератинов эпителиальных клеток, которые чаще вовлекаются в основные морфогенетические процессы развития и возникновения злокачественных опухолей (когда отсутствие апоптоза способствует прогрессированию заболевания).
Ламины и кератины типа I, в линкере, в середине а-спирализованного стержневого домена, содержат сайт чувствительности к каспазе 6. В кератинах типа II этот сайт отсутствует, однако поскольку они не способны полимеризоваться без участия кератинов типа I, достаточно удалить лишь один из двух типов кератинов, чтобы разрушить в клетке всю сеть промежуточных филаментов. Сайт чувствительности к каспазе существует также в С-концевом фрагменте белка К18. Этот белок атакуется каспазами 3 и 7 до момента проявления других признаков наступления апоптоза (таких как деградация ДНК или потеря поляризации мембраны). Таким образом, он представляет собой уникальный эпитоп, позволяющий детектировать ранние стадии апоптоза с помощью антител.