Механизм сегрегации хромосом у бактерий (прокариот)
• У прокариот отсутствует митотическое веретено, однако процесс сегрегации хромосом у них происходит безошибочно
• Исследование положения точек oriC на хромосоме показывает, что в раннем периоде цикла репликации ДНК они активно перемещаются к противоположным полюсам клетки
• Механизмы сегрегации хромосом выяснены недостаточно, вероятно, потому, что они отчасти являются функционально избыточными
• В процессе сегрегации хромосом у многих бактерий и низкокопийных плазмид вероятно, участвует система ParA-ParB
Подобно клеткам всех живых организмов, прокариоты должны обладать системами, гарантирующими, что до наступления деления вся генетическая информация полностью реплицировалась и что произошла точная сегрегация реплицированных пар сестринских молекул ДНК, по одной в каждую клетку. Исследования клеток бактерий дикого типа показали, что сегрегация представляет собой весьма эффективный процесс, и бесхромосомные клетки образуются с крайне низкой частотой, с трудом поддающейся регистрации (<10-4 в расчете на клетку в генерации).
В отличие от эукариот, у прокариот отсутствует явная структура митотического веретена, которая обеспечивает сегрегацию реплицированных хромосом. По-видимому, небольшие размеры прокариотических клеток исключают для них возможность образовывать веретено. Однако даже у эукариот дуплицированные центромеры должны быть пространственно разделены, с тем чтобы обеспечить их биполярное присоединение к веретену. Не исключено, что сегрегация хромосом у прокариот должна представлять собой аналогичный, или даже гомологичный, процесс по отношению к раннему, недостаточно исследованному этапу сегрегации хромосом у эукариот.
В ранних моделях сегрегации бактериальных хромосом предполагалось, что новореплицированная хромосома через область, содержащую oriC, присоединяется к клеточной оболочке с любой стороны центральной зоны роста. Затем элонгация клетки должна привести к тому, что параллельно с ростом клетки область сестринских oriС будет медленно отходить в сторону. В дальнейшем, однако, было показано, что рост клеточной оболочки не происходит по зональному принципу. Более того, в некоторых случаях процесс сегрегации хромосом осуществляется на больших расстояниях, как это, например, происходит при полярном делении спо-рулирующих клеток В. subtilis или через удлиненные палочки, как у некоторых стебельковых бактерий.
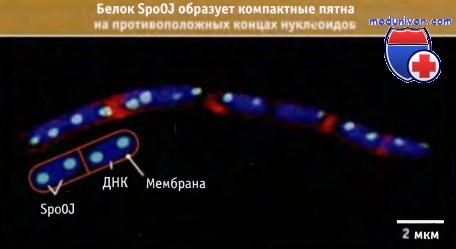
Визуализация белков SpoOJ, образующих компактные пятна в областях хромосом В. subtilis, содержащих oriС.
ДНК флуоресцирует голубым цветом (окраска DAPI), мембранные белки — красным (окраска FM5-95) и SpoOJ — зеленым (клетки экспрессируют конъюгат SpoOJ и зеленого флуоресцирующего белка).
На рисунке схематически представлена организация клетки, дуплицированные сайты oriC/SpoOJ расположены у внешних границ каждого нуклеоида.
Одна из причин, затрудняющих понимание процесса сегрегации хромосом у бактерий, заключается в трудности получения клеточных мутантов с нарушенной функцией сегрегации. У большинства выделенных мутантов функция сегрегации хромосом была затронута косвенным образом, а основные нарушения касались или процесса репликации, или общей организации нуклеоида. Это хорошо иллюстрируется на примере системы SMC (MukB). Путем генетического скрининга мутантов, образующих бесхромосомные клетки, были изолированы мутанты mukB. Оказалось, однако, что основная функция MukB и связанных с ним белков заключается в поддержании структурной организации нуклеоида (конденсации хромосом), хотя нельзя полностью исключить их участие в сегрегации. В последние годы в решении проблемы сегрегации хромосом у бактерий оказались плодотворными два подхода:
• Исследование сегрегации плазмид.
• Использование методов микроскопии для прямых наблюдений локализации и движения специфических сайтов хромосом при прохождении клетки по циклу.
Существует много механизмов, которые позволяют плазмидам находиться в стабильном состоянии. Репликация низкокопийных плазмид происходит таким образом, что количество их копий на клетку поддерживается близким к числу хромосом клетки хозяина (1-2 копии на клетку). Для плазмид характерны те же проблемы, связанные с декатенацией и разделением димеров, что и для хромосом. Обычно для преодоления всех этих проблем плазмиды используют такие системы клетки хозяина, как XerCD рекомбиназу. Многие плазмиды кодируют интересную систему, которая называется «яд — антидот» и использует другой подход для решения проблемы поддержки своей стабильности. Эта система действует таким образом, что дочерние клетки, не получившие копии плазмид, выбраковываются. В конечном счете, большинство низкокопийных плазмид приобрели активный механизм сегрегации, включающий два белка с общими названиями ParA и ParB.
Белки ParA обладают слабой АТФазной активностью и часто проявляют дополнительные функции в качестве регуляторов транскрипции. ParB относятся к категории белков, связывающихся с ДНК, и способны связываться со специфическими цис-действующими сайтами, необходимыми для сегрегации. Белок ParA может взаимодействовать с ParB, связанным со своим сайтом сегрегации, и поэтому также необходим для сегрегации. К сожалению, несмотря на почти 20-летние исследования, механизм, посредством которого в клетках прокариот обеспечивается стабильная сегрегация хромосом, остается неясным. Оказалось, что у большинства бактерий присутствуют гомологи белков ParA и ParB, необходимые для сегрегации хромосом (впрочем, у Е. coli и родственных бактерий они не обнаружены!). У В. subtilis в сегрегации и споруляции участвуют гомологичные белки, Soj и SpoOJ соответственно. Хотя хромосомы у этих бактерий часто показывают аномальное расположение, нулевые мутанты spoOJ жизнеспособны, что подтверждает точку зрения о том, что сегреграция хромосом является процессом, характеризующимся избыточностью. Белок SpoOJ связывается с несколькими сайтами, находящимися в протяженной области, порядка 800 кнп, расположенной поблизости от oriC.
Как показано на рисунке ниже, связанные белки сконцентрированы в виде компактных пятен, которые можно наблюдать во флуоресцентном микроскопе. Такая конденсация происходит с участием белка Soj. Так же как и для белков ParAB, механизм сегрегации хромосом с участием Soj остается неизвестным. Однако исследования пятен SpoOJ показывают, что вскоре после завершения раунда репликации ДНК наблюдается активная сегрегация области oriС. Пятна быстро занимают положение на противоположных концах реплицирующегося нуклеоида. К такому же выводу пришли на основании наблюдений над областью oriC, используя другие методы ее визуализации. Однако по меньшей мере в одном сообщении отмечается, что движение пятен происходит постепенно, что может свидетельствовать в пользу пассивной сегрегации. В настоящее время мы не можем представить себе ясную картину, описывающую участие белков системы ParAB в сегрегации хромосом. Действительно, оказалось, что в отсутствии этой системы быстро происходит начальный этап сегрегации областей хромосом, содержащих oriC. Высказано предположение о том, что движущей силой сегрегации может являться выход ДНК из фиксированной реплисомы.
Впрочем, такой механизм представляется маловероятным, поскольку репарация сестринских хромосом служит важным механизмом поддержки процессивности процесса их репликации. У многих типов бактерий в процессе активной сегрегации хромосом может участвовать белок MreB (актин), и эти данные представляют собой важное направление дальнейших исследований.