• Исследования, проведенные на разнообразных организмах, способствовали расширению наших знаний о регуляции клеточного цикла. Каждая из экспериментальных систем имеет свои преимущества и недостатки
• Исследования клеточного цикла дрожжей генетическими методами позволили идентифицировать консервативные регуляторы клеточного цикла
• Биохимический анализ белковых комплексов в клетках многоклеточных дополняет и расширяет генетические исследования, проведенные на одноклеточных
• Популяции синхронных клеток являются важной моделью для изучения процессов цикла
Перед тем как рассмотреть систему контроля клеточного цикла, нам следует охарактеризовать экспериментальные системы и модели, которые позволили исследователям прийти к важным заключениям. Эти системы крайне разнообразны: они варьируют от одноклеточных организмов до яйцеклеток амфибий и культур клеток человека. Каждая система обладает определенными преимуществами в исследованиях цикла, и в общем, те закономерности которые справедливы для одной из них, оказываются справедливыми для большинства других, поскольку основные механизмы регуляции клеточного цикла сохранились в ходе эволюции.
Клетки некоторых организмов, которые привлекли внимание исследователей, проявляют присущую им способность делиться синхронно. Например, при обработке соответствующими гормонами ооцитов морского ежа (Arbacia punctulata), шпорцевой лягушки (Xenopus laevis) и моллюска (Spisula salidissima) они вступают в синхронный цикл мейотического созревания. Гормональная стимуляция переводит ооцит из состояния интерфазы (незрелый ооцит) в состояние метафазы, в котором он готов к оплодотворению.
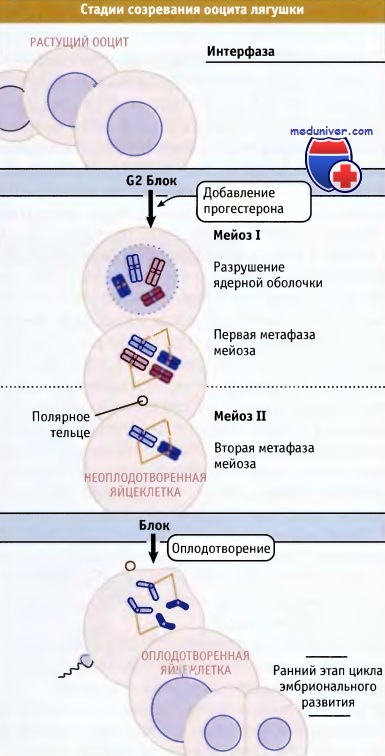
На рисунке ниже представлены стадии созревания и оплодотворение ооцита. Раннее деление эмбриональных клеток, наступающее после оплодотворения, также носит синхронный характер. Синхронное деление позволяет исследовать поведение клеточной популяции, а не индивидуальную клетку. Другое преимущество использования ооцитов или яйцеклеток (оплодотворенных ооцитов) моллюсков или амфибий для исследования клеточного цикла состоит в том, что эти объекты позволяют собрать большое количество материала, необходимого для биохимических анализов.
Крупный размер этих клеток позволяет проводить в них микроинъекции биологически важных молекул, например белков, а также химических препаратов, и таким образом изучать их влияние на процессы клеточного цикла. Из ооцитов и яйцеклеток готовят цитоплазматические экстракты, которые можно хранить для последующих экспериментов. Эти экстракты сохраняют способность к сборке вокруг ДНК ядерной оболочки, к репликации экзогенной ДНК, и к образованию митотического веретена в том же порядке, как эти процессы происходят в клеточном цикле. Таким образом, процессы клеточного цикла можно воспроизвести в пробирке. Кроме этого, клеточные экстракты можно использовать несколько раз. Такие свойства ооцитов моллюсков и амфибий широко использовались для очистки и идентификации белков, участвующих в процессах клеточного цикла, для изменения их внутриклеточного содержания и для проверки различных гипотез, относящихся к системе контроля цикла in vitro.
Для исследования клеточного цикла также широко используются клетки почкующихся и делящихся дрожжей (Saccharomyces cerevisiae и Schizosaccharomyces pombe). У этих одноклеточных эукариот осуществляются все основные процессы клеточного цикла, и они обладают многими преимуществами перед многоклеточными. Дрожжи легко выращивать в лабораторных условиях, и с ними просто работать. Они растут быстро: продолжительность цикла деления составляет 1-4 ч, легко поддаются простым методам синхронизации, например отбору клеток по размеру, а также генетическим манипуляциям. Одно из наиболее существенных отличий в клеточном цикле этих организмов и многоклеточных эукариот состоит в том, что в митозе ядерная оболочка дрожжевых клеток не разрушается; таким образом, у них происходит «закрытый митоз». Однако у дрожжей присутствует система контроля клеточного цикла и все контрольные точки.
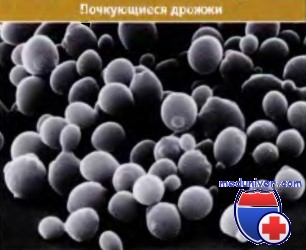
Клетки почкующихся дрожжей, как показывает их название, реплицируются, образуя почку, которая растет в интерфазе и в митозе отпочковывается от предсуществующей материнской клетки, образуя новую дочернюю клетку. Размеры почки указывают на стадию клеточного цикла, в которой находится клетка. Например, клетки, не содержащие почки, находятся в стадии G1, клетки с большой почкой — в стадии G2 или М. На рисунке ниже показана группа почкующихся клеток дрожжей.
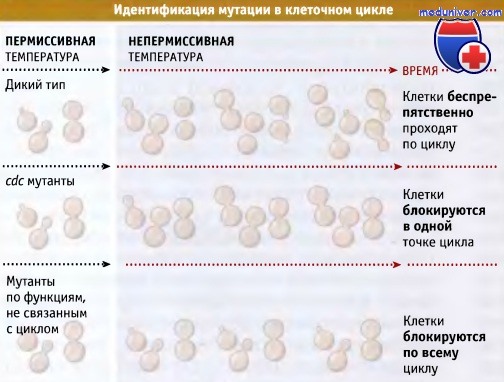
Как почкующиеся, так и делящиеся дрожжи могут расти как гаплоидные клетки и как условные мутанты с утратой функции. Если разработан соответствующий метод отбора, можно выделить мутанты, дефектные по любому процессу клеточного цикла. Хартвелл с сотрудниками выделяли мутанты, перенося клетки из условий роста при пермиссивной температуре 25 °С в непермиссивные условия (37 °С). При этом каждая колония мутантных клеток прекращает рост и погибает. У всех клеток почки оказываются одинаковой величины.
Блокирование роста на стадии образования однаковых почек указывает на то, что мутантные клетки дефектны по процессу перехода через специфическую стадию клеточного цикла. Например, мутанты, остановленные в росте и образующие почки большого размера, дефектны по процессу сегрегации хромосом. Температурочувствительные мутанты, выделенные Хартвеллом с сотрудниками, получили название cdc мутанты (англ, cell division cycle, cdc). Характеристика температурочувствительных cdc мутантов представлена на рисунке ниже.
Растущий ооцит остановлен в интерфазе. Прогестерон служит сигналом для его прохождения через мейоз I и вступления в мейоз II.
В ожидании оплодотворения ооцит блокируется в метафазе мейоза II.
При оплодотворении этот второй блок цикла снимается, и затем оплодотворенная яйцеклетка проходит по циклу эмбрионального развития.
Скрининг этих мутантов оказался крайне полезным при идентификации генов, регулирующих различные процессы клеточного цикла. Еще одна полезная особенность температуро-чувствительных мутантов состоит в том, что блокирование роста клеток на стадии формирования определенного фенотипа и обратимость некоторых мутаций (они не выходят из клеточного цикла при переключении от непермиссивных к пермиссивным условиям) служат хорошим способом синхронизации всей популяции таких клеток.
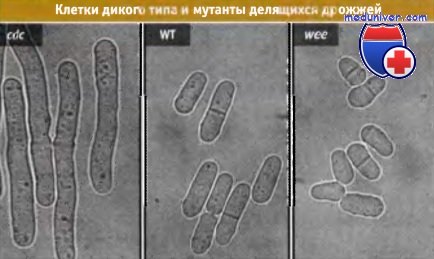
В отличие от почкующихся, клетки делящихся дрожжей обладают цилиндрической формой, растут за счет элонгации концов, и делятся с участием медиально расположенной перегородки. В популяции клеток дикого типа, растущих на богатой среде, длина клеток при делении постоянна, и поэтому длина клетки S. pombe служит хорошим показателем стадии клеточного цикла. Как и ожидалось, в экспериментах Нерза и сотрудников длина клеток S. pombe оказалась идеальным морфологическим маркером для визуальной идентификации cdc мутантов. Мутантные клетки не делились, но становились длиннее, чем клетки дикого типа. Этот признак также помог выделить «крошечные» («wee») мутанты, которые делились при уменьшении размеров клеток. Рисунок ниже иллюстрирует различия в длине между клетками дикого типа и мутантами делящихся дрожжей.
Выделение cdc и wee мутаций по одному генетическому локусу (cdc2+) позволило предположить, что в клетках существует важный фактор, регулирующий наступление митоза. Ниже в этой главе мы подробнее рассмотрим роль Cdc2.
Расширению наших знаний о процессах клеточного цикла способствовали эксперименты с использованием еще двух организмов. Эти организмы позволяют легко отбирать клетки, мутантные по процессам контроля клеточного цикла, и работать с ними. Это филаментозный гриб Aspergillus nidulans и плодовая мушка Drosophila melanogaster. Однако они используются реже дрожжей. У этих организмов клетки синхронно проходят через ранние клеточные циклы. Бесполые споры Anidulans находятся в G1 стадии клеточного цикла, и первые два или три цикла деления протекают с высокой степенью синхронизации. Наблюдения за клеточным циклом у Anidulans также позволили выявить его скоординированность с процессами развития, чего нельзя было сделать, используя дрожии в качестве объекта исследования.
Аналогичным образом, наблюдение за ранними делениями эмбриона мухи позволяет выявить синхронные циклы ядрного деления, которые происходят в общей цитоплазме. Такие возможности, а также прекрасно исследованная генетика и знание особенностей развития организма делает D. melanogaster ценным объектом для исследований скоординированности сигналов клеточного цикла с процессами развития.
Исследования на культурах клеток млекопитающих также способствовали существенному прогрессу в области изучения механизмов контроля процессов клеточного цикла. В идеале, было бы желательно иссследовать цикл на первичной культуре клеток млекопитающих, т. е. на клетках, которые непосредственно взяты из организма, и не подвергались генетическим изменениям. Однако первичная культура клеток не обладает способностью делиться неопределенно долго. Через 25-40 циклов они прекращают деление и начинают стареть (подробнее об этом см. ниже). Поэтому для анализа событий клеточного цикла широко используют иммортализованные линии, полученные из нормальных или раковых клеток.
Клетки почкующихся дрожжей в сканирующем электронном микроскопе.
Непочкующиеся клетки находятся в стадии G1, в то время как крупные почкующиеся клетки — в стадии G2 и М.
Как показывает само название, иммортализованные клетки являются генетически измененными таким образом, что обладают способностью к бесконечному делению в культуре, находясь в соответствующей питательной среде, содержащей ростовые факторы. Хотя всегда следует иметь в виду, что свойства таких клеток не всегда совпадают с соответствующими свойствами клеток организма, линии иммортали-зованных клеток, например HeLa, оказались чрезвычайно полезными объектами при изучении клеточного цикла Они легко поддаются синхронизации с помощью ингибиторов синтеза ДНК или митоза, и/или сортировке по размерам, а также могут быть использованы для цитологических и биохимических исследований.
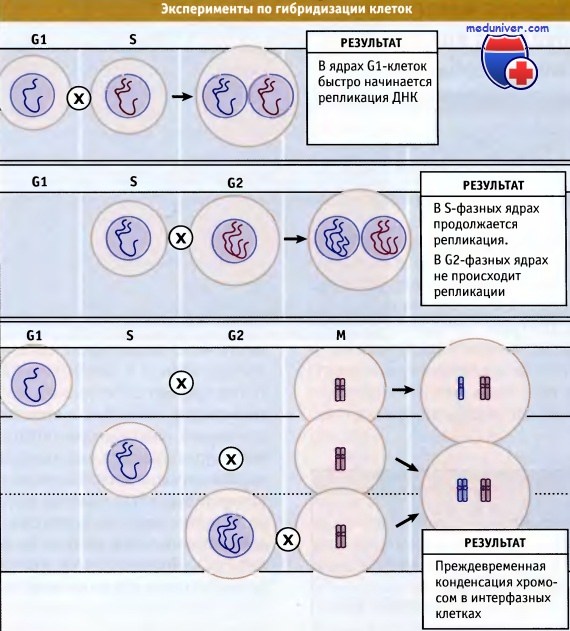
В самом деле, элегантные эксперименты по слиянию клеток млекопитающих заложили основу для представлений о контрольных точках и о скоординированности процессов клеточного цикла. В этих экспериментах слиянию друг с другом подвергали клетки, находящиеся на разных стадиях клеточного цикла (G1, S, G2 и М). Оказалось, что при слиянии S-фазных клеток с клетками, находящимися в фазе G1, ускоряется вступление ядер последних в фазу репликации ДНК Этот результат позволил высказать предположение о том, что промотирующие факторы S-клеток переносятся в гибридные G1/S-клетки через цитоплазму.
Важные данные относительно основных факторов, влияющих на прохождение клеток по циклу, были получены в экспериментах по слиянию митотических клеток с клетками, находящимися в фазах G1, S и G2. Во всех случаях в интерфазных клетках наблюдалась преждевременная конденсация хромосом, и клетки вступали в псевдомитоз (как бы вступали в митоз). Эти эксперименты предполагают существование индуктора митоза, играющего доминантную роль, поскольку он индуцирует конденсацию хромосом в ядрах клеток, находящихся на любой стадии клеточного цикла. Оказалось, что этот индуктор присутствует в клетках всех эукариот, и он был выделен и охарактеризован из многих типов клеток.
В этих экспериментах также было показано, что, когда С2-клетки сливались с S-фазными, С2-фазные ядра не проявляли ингибирующего действия на репликацию, происходящую в S-фазных ядрах. Однако G2-фазные ядра также не начинали репликации своей ДНК. Это наблюдение позволило высказать предположение о «лицензировании» ДНК, согласно которому репликация ДНК происходит только один раз за цикл. Схемы экспериментов по слиянию клеток представлены на рисунке ниже.
Наряду с установлением основных принципов автономной регуляции клеточного цикла, культуры клеточных линий позволяют исследовать влияние на него внешних стимулов. Это обеспечивает разработку вопросов, связанных с регуляцией роста и деления клеток млекопитающих генами — супрессорами опухоли и онкогенами (подробнее см.
Наконец, для выяснения функции генов клеточного цикла у млекопитающих можно использовать мышей с выключенными функциями определенных генов. Эти эксперименты позволяют выяснить, необходимы белки клеточного цикла д ля всех клеток или только для развития и функционирования клеток некоторых тканей. Возможно также определить, насколько потеря какого-либо белка клеточного цикла у млекопитающих может быть скомпенсирована другим белком и насколько более склонными к развитию таких заболеваний, как рак, являются животные, у которых отсутствует регулятор клеточного цикла.
Каждая экспериментальная система имеет свои преимущества и свои недостатки. Например, пригодность ооцитов лягушки, моллюска и морского ежа для биохимических исследований дает им преимущество перед другими объектами при проведении экспериментов по реконструкции клеточных процессов in vitro. В то же время с помощью хорошо разработанной генетики дрожжей, грибов и плодовой мушки был проложен путь к идентификации ключевых регуляторов прохождения клеток по циклу. При использовании всех экспериментальных систем был получен колоссальный объем информации, касающейся механизмов контроля клеточного цикла.
При росте при пермиссивной температуре 25 °С почкующиеся дрожжи S. cerevisiae дают смесь непочкующихся, мелких и крупных почкующихся клеток.
Однако если, например, в гене, регулирующем митоз, присутствует температуро-чувствительная мутация,
то в условиях непермиссивной температуры 37 °С, дрожжи неизменно останавливаются в росте в виде крупных почкующихся клеток.
cdc мутанты не делятся и образуют более длинные клетки, чем дрожжи дикого типа.
У мутантов wee деление наступает раньше, чем у дрожжей дикого типа (WT), что приводит к появлению мелких клеток.
Существование таких фенотипов указывает на наличие мутаций в системе регуляции цикла.
Фотографии представляют собой изображения cdc мутантов (слева), клеток дикого типа (в середине),
и wee мутантов S pombe (справа), полученные с помощью DIC-микроскопии.
(Верхний ряд) Когда клетки млекопитающих, находящиеся в G1-фазе, гибридизуют-ся с S-фазными клетками, ядра G1-клеток быстро начинают реплицировать ДНК.
(Средний ряд) Когда S-фазные клетки гибридизуются с находящимися в G2-фазе, то в последних не происходит повторной репликации ДНК,
и также не подавляется репликация, идущая в S-фазных клетках. Они также не вступают в митоз.
Скорее, до вступления в митоз, клетки ожидают завершения репликации в S-фазных ядрах.
(Нижний ряд) Когда интерфазные клетки млекопитающих (G1, S или G2) гибридизуются с митотическими клетками,
в интерфазных клетках сразу же начинается конденсация хромосом, и они вступают в псевдомитотическое состояние.
Это свидетельствует о том, что митотические клетки содержат доминантный фактор, способствующий митозу.
Учебное видео митоз и клеточный цикл
Скачать данное видео и просмотреть с другого видеохостинга можно на странице: Здесь.