• В митозе веретено прикрепляется к кинетохорам хромосом
• Необходимым условием сегрегации хросомом служит правильное прикрепление микротрубочек к кинетохорам
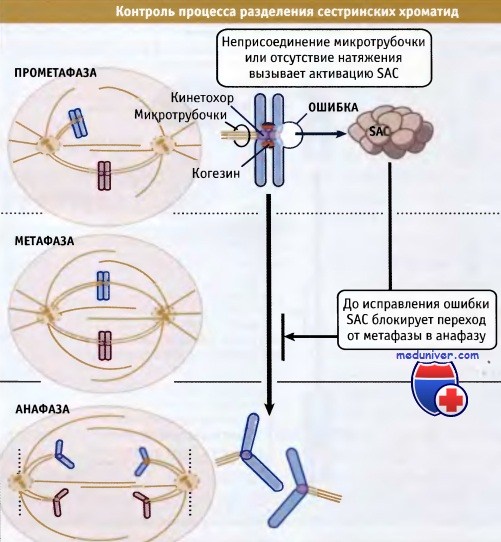
• Дефекты прикрепления микротрубочек веретена узнаются «точкой контроля за сборкой веретена», которая в дальнейшем блокирует переход из метафазы в анафазу, чтобы не допустить ошибок при разделении сестринских хроматид
На ранних стадиях митоза микротрубочки веретена (МТ) прикрепляются к кинетохорам, представляющим собой белковые комплексы, которые собираются на центромерных участках ДНК хромосом. Для правильной сегрегации хромосом в дочерние клетки необходимо, чтобы до начала анафазы кинетохоры сестринских хроматид были прикрепленными к МТ, исходящим из противоположных полюсов веретена. Такие прикрепление называется амфителическим или биполярным.
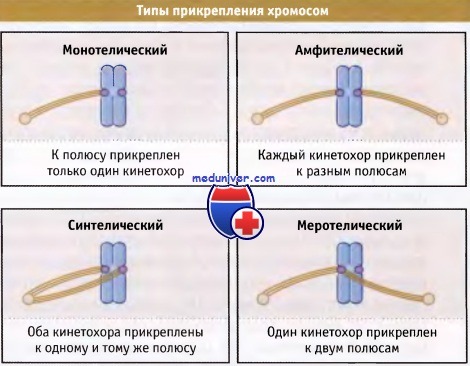
Поскольку прикрепление кинетохора к микротрубочке веретена (МТ) в метафазе происходит случайным образом, возможно образование и других типов прикреплений. На рисунке ниже представлены разные типы прикреплений кинетохоров к МТ.
В клетке существуют механизмы, позволяющие отличить правильные (биполярные) прикрепления от неправильных (монополярных). Несколько белков участвуют в метафазе в превращении неправильных прикреплений кинетохора к МТ в правильные, амфителические. Ключевым белков, дестабилизирующим неправильное прикрепление, является протеинкиназа, Aurora В.
Как отмечалось выше, эта киназа является компонентом комплекса белков-пассажиров хромосом и в митозе локализована на кинетохорах. Механизм функционирования киназы Aurora В в обеспечении биполярного прикрепления хромосом к веретену служит предметом интенсивных исследований.
Что происходит, когда некоторые хромосомы амфителически не прикрепляются к веретену или вообще не прикрепляются? В этих случаях «точка контроля за сборкой веретена» (SAC) генерирует сигнал «ждать анафазу». Эта точка контроля задерживает разделение сестринских хроматид, т. е. наступление анафазы до тех пор, пока каждый кинетохор правильно не прикрепится к микротрубочкам веретена.
В монотелической конфигурации только один кинетохор (выделен пурпурным цветом) прикреплен к фибриллам веретена, исходящим от одного полюса.
В амфителической конфигурации кинетохоры сестринских хроматид связаны с микротубочками веретена от противоположных полюсов, что приводит к правильной биполярной ориентации хромосомы.
В синтелической конфигурации оба кинетохора сестринских хроматид прикреплены к микротрубочкам веретена, идущим от одного полюса.
В меротелической конфигурации один кинетохор прикреплен к микротрубочкам веретена от обоих полюсов.
Все неправильные конфигурации узнаются контрольной точкой сборки веретена, в результате чего митоз блокируется.
Если сигнал «ждать анафазу» не генерируется, разделение сестринских хроматид последует раньше времени, что приведет к сегрегации неравного числа хромосом. SAC блокирует наступление анафазы, предотвращая активацию АРС и тем самым деградацию секурина.
Как происходит активация SAC? Процесс инициируется кинетохорами, не прикрепленными или неправильно прикрепленными к МТ веретена. Хотя показано, что неприкрепленные кинетохоры индуцируют активацию SAC, существуют данные о том, что в отсутствие на каждом кинетохоре физических напряжений активируется точка контроля за сборкой веретена. Прежде чем стали известны молекулярные механизмы функционирования SAC, активацию этой точки исследовали на сперматоцитах кузнечика. В клетках кузнечика присутствуют три половых хромосомы, и в некоторых клетках одна из них остается неспаренной и прикрепляется только к одному полюсу веретена.
Клетки, содержащие неприкрепленный кинетохор, никогда не вступают в анафазу и в конце концов разрушаются. В изящных экспериментах с помощью микроиглы пытались оттянуть неприкрепившуюся хромосому. При этом клетка вступала в анафазу, что говорило о том, что для снятия эффекта SAC также необходимо наличие на кинетохоре напряжения.
Как можно ожидать, вследствие биполярного прикрепления микротрубочки веретена (МТ) к сестринским кинетохорам на них развивается напряжение. В то же время при других типах прикреплений такое напряжение не создается. В настоящее время предложена модель, которая в качестве важных параметров мониторинга SAC учитывает прикрепление кинетохоров и создание на них физического напряжения.
Впервые SAC и ее компоненты были идентифицированы при генетическом скрининге мутантов почкующихся дрожжей, чувствительных к соединениям, разрушающим микротрубочки. Однако эта точка также существует у многоклеточных эукариот. Интересно, что компоненты SAC не обязательны для вегетативного роста дрожжей, но в экспериментах на мышах показано, что они необходимы для обеспечения жизнеспособности эмбрионов. Клетки, полученные от эмбрионов, лишенных некоторых компонентов SAC, обнаруживают повышенную частоту ошибок сегрегации хромосом.
Необходимость в SAC у Метазоа, вероятно, обусловлена большей длительностью у них процесса выравнивания хромосом и прикрепления кинетохоров к МТ, в течение которого АРС должен находиться в неактивном состоянии. Функционирование SAC схематическти показано на рисунке ниже.
Каким образом SAC задерживает наступление анафазы? Предстоит еще многое выяснить в отношении механизмов узнавания дефектов этой контрольной точкой и передачи соответствующего сигнала. Однако очевидно, что сигнал активации SAC распространяется от кинетохоров. В них обнаружены все компоненты процессов SAC. Это характерно для нормально протекающего клеточного цикла и для активации точки контроля. Вероятно, исходной точкой приложения компонентов точки контроля являются кинетохоры.
Неприсоединение микротрубочки к одному кинетохору или отсутствие надлежащего натяжения между сестринскими кинетохорами приводит к активации контрольной точки сборки веретена (SAC).
В результате, при переходе из метафазы в анафазу, митоз блокируется до момента устранения ошибки.
Более того, показано, что локализация компонентов в кинетохорах необходима для генерации и распространения сигнала контрольной точки. Процесс накопления компонентов SAC в кинетохорах нарушается при некоторых дефектах структуры последних, связанных с мутациями по генам соответствующих белков. У таких мутантов не функционирует точка контроля. Прочие дефекты структуры кинетохоров не препятствуют активации SAC.
Известно, что система сигналов SAC включает каскад фосфорилирования белков, поскольку некоторые ее компоненты представляют собой протеинкиназы, а многие находятся в фосфорилированной форме. Однако накапливается все больше данных о том, что передача сигнала не носит строго линейного характера. Например, для мониторинга различных аспектов биполярного прикрепления необходимы различные компоненты. Неизвестно также, существуют ли множественные мишени для протеинкиназ системы SAC.
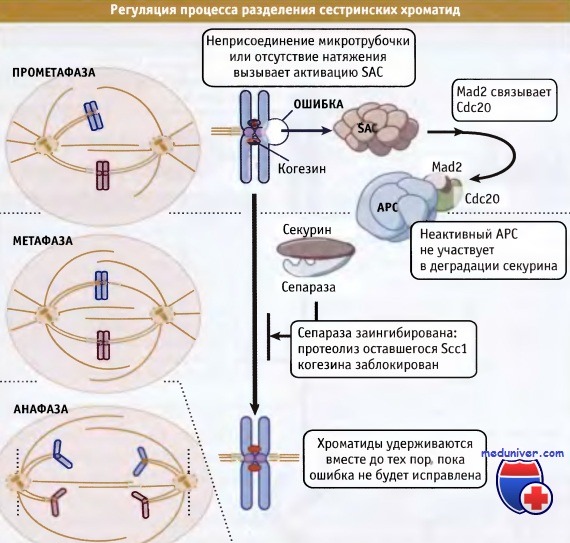
Одной из мишений является белок Cdc20. Как отмечалось выше, Cdc20 представляет собой один из специфических активаторов АРС. Также Cdc20 связывает другой белок системы SAC, Mad2. Это позволяет предполагать, что последний может ингибировать АРС, вытесняя из комплекса с ним Cdc20. Недавно был идентифицирован комплекс контрольной точки митоза — MCC (mitotic checkpoint complex), содержащий три белка — Mad2, BubR1, Bub3, а также Cdc20. Хотя неизвестно, один ли Mad2, или вместе с МСС, генерируют сигнал задержки наступления анафазы, очевидно, что после активации точки контроля APCCdc20 активируется, что стабилизирует секурин и поддерживает связь между сестринскими хроматидами. Модель, описывающая ингибирование АРС под действием SAC, представлена на рисунке ниже.
Основная функция SAC состоит в предотвращении сегрегации хромосом при неправильном прикреплении сестринских кинетохоров к микротрубочкам веретена. Как можно ожидать, функционально неактивная SAC привела бы к дефектам прикрепления кинетохоров, неправильной сегрегации хромосом и к гибели клетки, или к появлению их анеуплоидных форм. Дочерние анеуплоидные клетки получают неравные количества генетического материала. Анеуплоидия характерна для многих форм злокачественных опухолей, и мутации по отдельным компонентам SAC связаны с возникновением некоторых форм колоректальных раков.
Убедительные доказательства участия SAC в предотвращении повреждения хромосом в митозе были получены в экспериментах по снижению уровня Mad2 в клетках мышей за счет элиминации одной копии хромосом. У животных с таким дефектом часто развивались злокачественные опухоли, а клетки со сниженным уровнем Mad2 обнаруживали повышенное число ошибок сегрегации хромосом.
Суммируя вышесказанное, подчеркнем, что функционирование точки контроля за сборкой веретена гарантирует, что сегрегация хромосом произойдет только после того, как все хромосомы правильно прикрепятся к митотическому веретену. Поскольку сегрегация хромосом инициируется активацией АРС, точка контроля не допускает этой активации до момента правильного прикрепления всех хромосом. Сигнал, который подает контрольная точка генерируется на кинетохоре, и приводит к прямому связыванию некоторых ее компонентов с АРС, что вызывает ингибирование функций комплекса.
При активации контрольной точки сборки веретена (SAC) происходит связывание Mad2,
одного из компонентов SAC, или комплекса его компонентов (МСС) с Cdc20, который является активатором АРС.
Это связывание приводит к ингибированию активности АРС Cdc20.
Последний необходим для протеолиза секурина и обеспечения расхождения сестринских хроматид.