• Существует точка контроля, предотвращающая наступление анафазы до того момента, как все кинетохоры прикрепятся к митотическому веретену
• Неприкрепившиеся кинетохоры продуцируют сигнал, который не позволяет анафазе начаться
• Точка контроля проверяет количество микротрубочек, прикрепленных к кинетохору
• Когда все кинетохоры в клетке правильно прикрепились к веретену, происходит активация анафазного промоторного комплекса (АРС)
• Активация АРС приводит к деструкции белков, удерживающих сестринские хроматиды вместе
Расхождение хроматид служит визуальным показателем завершения клеткой метафазы и перехода ее в анафазу. Наряду с разрушением оболочки ядра, это наиболее выраженное видимое изменение, происходящее на протяжении жизненного цикла клетки. Так же как и разрушение ядерной оболочки, расхождение хроматид представляет собой необратимый процесс.
Если расхождение сестринских хроматид происходит до момента прикрепления всех хромосом к обоим полюсам веретена, то часто клетка становится анеуплоидной. Для предотвращения этого в клетке существует точка контроля, в которой проверяется прикрепление кинетохоров к веретену при митозе. Неприкрепленные или слабо прикрепленные кинетохоры, которые могут свидетельствовать о неполноте процесса сборки веретена, продуцируют сигнал, который задерживает наступление анафазы до момента исправления ошибок, — сигнал «ожидания анафазы».
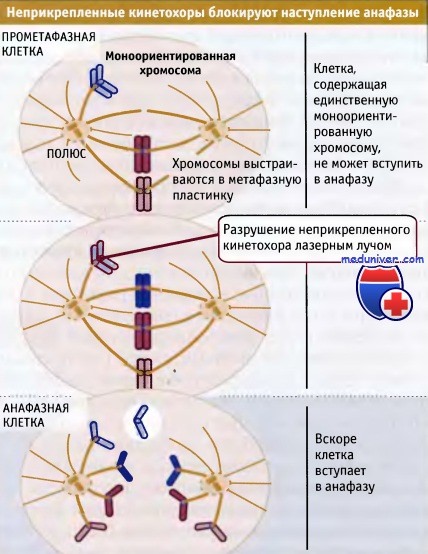
Мы знаем, что такой сигнал существует, поскольку обычно анафаза не наступает, пока клетка содержит хотя бы одну хромосому в моноориентации. Однако анафаза наступает сразу, если с помощью лазерного микропучка разрушить неприкрепившийся кинетохор этой хромосомы. Сигнал продуцирутся автоматически, как только присходит разрушение ядерной оболочки, поскольку кинетохоры высвободившихся хромосом все оказываются неприкрепленными. Поэтому сигнал постоянно присутствует до тех пор, пока последний кинетохор не прикрепится к веретену.
Эта точка называется точкой контроля прикрепления кинетохоров, или точкой контроля за сборкой веретена. Она обеспечивает гарантию того, что в подавляющем большинстве случаев произошла правильная сегрегация хромосом. Однако при этом не обеспечивается контроль над возможностью ошибочного прикрепления одного кинетохора к двум полюсам (т. е. в случае меротелической ориентации).
Невозможность обнаружения этого дефекта означает, что иногда анафаза наступает до того, как ошибка будет исправлена. Хотя ошибки сегрегации хромосом, как правило, редки, они все-таки возникают с частотой одна ошибка на каждые 10 000 делений. Имеющиеся данные позволяют предполагать, что меротелическое прикрепление является основной причиной образования анеуплоидных клеток.
Схема эксперимента, иллюстрирующего, что неприкрепленный кинетохор генерирует сигнал, который предотвращает наступление анафазы.
Наверху — клетка, остановленная в прометафазе. Вскоре после инактивации компонентов неприкрепившегося кинетохора с помощью ультратонкого лазерного луча наступает анафаза.
В точке контроля сборки веретена проверяется, сколько микротрубочек прикрепилось к кинетохору. Если их слишком мало, то кинетохор генерирует сигнал ожидания анафазы. Этот сигнал выключается, когда количество микротрубочек, прикрепленных к кинетохору, превышает критическую величину. Поскольку натяжение на кинетохоре стабилизирует его и способствует накоплению микротрубочек, точка контроля оказывается чувствительной к натяжению на кинетохоре. Максимальное натяжение развивается, когда сестринские кинетохоры присоединяются к противоположным полюсам, так что при проверке количества микротрубочек на кинетохорах в точке контроля косвенным образом оценивается, насколько правильно прикреплены к веретену все хромосомы.
При разрушении микротрубочек веретена с помощью химических соединений, точка контроля сборки веретена остается активной в течение продолжительного времени. Постоянное присутствие сигнала при воздействии на клетку таких соединений было использовано для отбора мутантов дрожжей, дефектных по генам, необходимым для функционирования точки контроля. Были идентифицированы гены, кодирующие три белка Mad (mitosis arrest deficient) и три белка Bub (budding uninhibited by benzamidazole). У этих белков есть гомологи в клетках позвоночных и человека. Если блокировать активность этих белков, то в обход точки контроля быстро наступает анафаза.
В митозе некоторые из этих белков, включая BubR1 и Mad2, постоянно то появляются на неприкрепленных кинетохорах, то отщепляются от них, но не обнаруживаются на прикрепленных кинетохорах.
Критическим событием, которое инициирует анафазу, обеспечивающую расхождение сестринских хроматид, является активация большого макромолекулярного комплекса, известного как анафазный промоторный комплекс. Функция этого комплекса заключается в отборе белков, которые необходимо разрушить, и это достигается путем добавления к ним молекул убиквитина. Последний играет роль маркера, который позволяет протеолитическим системам клетки (протеосомам) узнавать белок и деградировать его. Однако анафазный промоторный комплекс не может функционировать в изолированном виде.
Для него необходим кофактор, определяющий, какие белки необходимо разрушить и в какое время. Наступление анафазы предполагает, что анафазный промоторный комплекс активируется кофактором Cdc20. В точке контроля сборки веретена происходит ингибирование активности Cdc20, которая, в свою очередь, не позволяет комплексу узнавать белок секурин с тем, чтобы не допустить его разрушения. Разрушение секурина приводит к деструкции склеивающих хромосомальных белков, которые скрепляют реплицированные хроматиды.
Каким образом кинетохор и компоненты механизма точки контроля влияют на белок Cdc20, пока неизвестно. Вопрос заключается в том, каким образом сигнал передается от кинетохора на веретено, в котором находится анафазный промоторный комплекс. Согласно одной точке зрения, на кинетохоре образуется комплекс, состоящий из белков Mad2, BubR1, и Cdc20, который затем высвобождается.
Другая возможность заключается в том, что неприкрепленные кинетохоры связываются и активируют один или несколько белков, участвующих в процессах контроля, а затем высвобождают активированные белки в область веретена, где они образуют комплексы с Cdc20, который предотвращает их от активности АРС. Хотя неизвестно, где образуются комплексы между Cdc20 и компонентами системы контроля, очевидно, что они образуются постоянно, пока существуют неприкрепившиеся кинетохоры.
Однако комплексы существуют недолго, так что при прикреплении последнего кинетохора ингибирующий эффект более не проявляется. Каким образом сигнал ожидания анафазы, который подается последним неприкрепившимся кинетохором, преобразуется в доступную для всего веретена форму, еще предстоит выяснить.