Механизм конденсации и сегрегации хромосом в митозе
• При подготовке к расхождению хромосомы конденсируются и мигрируют к центру митотического веретена
• Хромосомы прикрепляются к микротрубочкам, выходящим из противоположных полюсов веретена, с помощью специальных участков, называемых кинетохорами
• Разрываются связи, соединяющие сестринские хроматиды вместе, и происходит их разделение
• Происходит расхождение изолированных сестринских хроматид, что предшествует цитокинезу
Как отмечалось выше, у большинства клеток одним из характерных признаков вступления в митоз является конденсация хромосом. Генетический материал интерфазных клеток упакован не так плотно как в митозе, и в процессе сегрегации хромосом важную роль играет его реорганизация с образованием компактных структур высшего порядка. Как достигается эта компактизация и каким образом она регулируется?
В конденсации хромосом участвует мультибелковый комплекс, который называется конденсин. Этот комплекс состоит из двух суперспирализованных белковых молекул, содержащих АТФазные домены и, в силу исторических причин, относящихся к семейству SMC-белков. Предполагается, что субъединицы конденсина могут «прогуливаться» по ДНК, скрепляя вместе ее различные участки. В состав конденсинового комплекса также входят другие белки, которые, наряду с SMC белками, вызывают в ДНК конформационные и топологические изменения, приводящие к компактизации.
Доступ конденсинового комплекса к хромосоме тщательно контролируется. Комплекс связывается с хромосомой только во время митоза. В клетках сливающихся дрожжей, у которых не разрушается ядерная оболочка, конденсин физически отделен от хромосом, будучи заключен в пределах цитоплазмы. Фосфорилирование одной из субъединиц конденсина под действием Cdk1, происходящее в митозе, вызывает миграцию комплекса в ядро. У Xenopus Cdk1, вероятно, активирует процесс суперспирализации, происходящий при действии конденсина.
О процессе конденсации хромосом также говорят и другие изменения связанных с ними белков, например фосфорилирование гистонов Н1 и Н3. Известно, что фосфорилирование гистона Н3 происходит с участием киназ Aurora. Фактически, фосфорилирование этого гистона в митозе обычно служит показателем активности киназы Aurora.
Образование связей между сестринскими хроматидами представляет собой процесс, родственный конденсации хромосом. При репликации в S-фазе каждая пара хроматид связывается между собой и находится в связанном состоянии до момента разделения при переходе от метафазы к анафазе. Каким образом и когда устанавливаются связи между хроматидами и как они разрываются в анафазе?
Так же как и конденсиновый комплекс, когезиновый комплекс формирует и поддерживает связи между сестринскими хроматидами, начиная от момента репликации ДНК и до начала их сегрегации. Этот комплекс состоит из двух SMC-белков, которые напоминают SMC-белки конденсина, хотя и отличаются от них, и трех дополнительных белков, Scc1, Scc3 и Pds5, также необходимых для его функционирования.
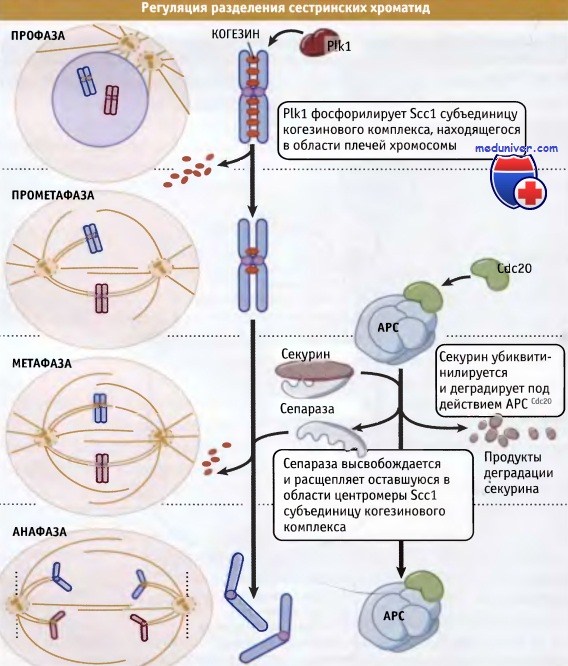
До наступления метафазы одна из протеаз, под названием сепараза, находится в связанном с секурином состоянии и не проявляет активности.
В метафазе, при действии АРС, секурин подвергается протеолизу, и таким образом происходит деблокирование сепаразы.
После этого активная сепараза расщепляет когезин, что вызывает разделение сестринских хроматид.
Предполагалось, что когезиновый комплекс скрепляет сестринские хроматиды наподобие застежки «молния». Однако недавно было показано, что SMC-белки и субъединица Scc1 обвивают сестринские хроматиды, скрепляя их. Подробности механизма этого скрепления сейчас исследуются.
Предложено два механизма разрушения связей между хроматидами. Согласно одному механизму, в профазе, с плечей хромосомы удаляется большая часть когезина, а связанный с центромерой комплекс остается на месте. Этот первый этап удаления когезина запускается Plk1, которая фосфорилирует Scc1. В то же время связанная с центромерой Scc1 когезиновая субъединица подвергается протеолизу. Если модель скрепления хроматид за счет их обвивания субъединицей Scc1 правильна, то расщепление этой субъединицы должно приводить к раскрытию обвивающего кольца и к физическому разделению сестринских хроматид. Во всяком случае, каким бы ни был механизм скрепления хроматид, очевидно, что разрушение Scc1 вызывает их диссоциацию и обеспечивает наступление анафазы.
Сепараза представляет собой сайт-специфическую протеазу, которая расщепляет Scc1 и вызывает разделение хроматид. В течение большей части клеточного цикла, сепараза поддерживается в неактивном состоянии, находясь в комплексе с другим белком, секурином. При переходе из метафазы в анафазу секурин подвергается убиквитинилированию и атакуется протеазами. При разрушении секурина высвобождается сепараза (также известная под названием сепарин), которая разрушает Scc1.
Процессы, происходящие при разделении хроматид, представлены на рисунке ниже. Таким образом, в этой простой модели ключевым процессом, запускающим сегрегацию хромосом, является протеолиз секурина. В связи с этим возникает интересный вопрос: каким образом регулируется деградация секурина?
Секурин отбирается для деградации при его взаимодействии с ЕЗ-убиквитинлигазой, известной под названием анафазный промоторный комплекс (АРС). Этот комплекс узнает белки, содержащие короткие последовательности, называемые деструктивными и/или KEN доменами (D boxes или KEN boxes). При добавлении этих последовательностей, содержащих около девяти аминокислот, в структуру белков, последние проибретают способность к деградации под действием АРС.
В отличие от SCF, для того чтобы субстрат был узнан АРС, не требуется его предварительного фосфорилирования. У почкующихся дрожжей существенная функция АРС состоит в деградации циклинов и секурина, хотя для этого комплекса известно также много других мишеней.
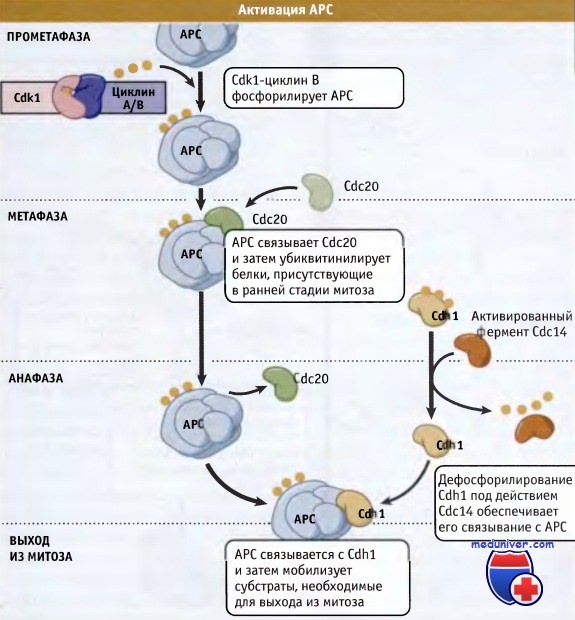
АРС проявляет активность только в митозе и G1-периоде, когда он предотвращает накопление циклинов и секурина. Как регулируется активность АРС? Во-первых, связывание адаптерных белков (Cdc20 и Cdh1 — обозначаемых как APCcdc20 APCcdh1) с APC обеспечивает значительную степень субстратной специфичности комплекса. Адаптерные белки способны связываться с АРС лишь в митозе и G1-периоде цикла. Во-вторых, активность АРС регулируется при специфическом фосфорилировании некоторых из его 13 субъединиц и адаптерных белков.
В фосфорилировании и активации АРС участвуют Cdkl и Plk1. В-третьих, активность АРС может контролироваться сигнальной системой, которая следит за прикреплением хромосом, а также точкой, контролирующей сборку веретена (SAC). Столь многочисленные уровни регуляции гарантируют, что АРС активируется только в митозе и запускает протеолиз секурина и циклина, активирует сепаразу, расщепляет когезин, и, наконец, вызывает разделение сестринских хроматид. В G1-периоде АРС сохраняет свою активность с тем, чтобы поддерживать активность Cdk1 на низком уровне. Низкая активность этой киназы позволяет сформироваться pre-RC, необходимому для следующего раунда репликации ДНК. Схема процессов активации АРС представлена на рисунке ниже.
Когда после разрыва связи сестринские хроматиды отделились друг от друга, они расходятся к противоположным полюсам клетки под действием микротрубочек веретена. Когда хромосомы разошлись и активность Cdk1 снизилась, наступает цитокинез.
Подводя итоги изложенному выше, подчеркнем, что конденсация хромосом происходит с участием белкового комплекса, который называется конденсин, и что сестринские хроматиды удерживаются вместе когезиновым комплексом до тех пор, пока не произойдет их разделение в анафазе. Разрыв связей между хроматидами требует расщепления когезина под действием протеазы, сепаразы. Для активации сепаразы необходим протеолиз ее ингибитора, секурина, который происходит с участием АРС по убиквитиновому механизму.
В интерфазе АРС убиквитин лигаза неактивна.
При наступлении митоза Cdk1 фосфорилирует АРС и он связывается с активатором, Cdc20.
Затем Cdc20 АРС узнает субстраты, например секурин, обеспечивая необратимое прохождение клетки по фазам митоза.
Позже в митозе АРС связывается с активатором Cdh1 и мобилизует другие субстраты, необходимые для выхода из митоза.
Первый видеокадр, показывающий хромосомы в начальных стадиях митоза.