• Клетки могут находиться в неделящемся состоянии, которое называется состоянием покоя или G0
• Покоящиеся клетки могут возвращаться в клеточный цикл под влиянием сигнала извне
• Клетки повторно входят в цикл в основном на стадии G1
• При дифференцировке в специализированные клетки, они также могут навсегда выйти из цикла
• Некоторые клетки запрограммированы на самоуничтожение путем апоптоза
Мы ввели понятие основной биологической машины клеточного цикла, которая осуществляет все события, необходимые для прохождения этого процесса. Однако эта машина «включена» не всегда, поскольку переход к делению, при отсутствии соответствующих внешних условий, представляет опасность для клетки. В данном разделе мы рассмотрим, как информация, извне поступающая в клетку, преобразуется в решение, включать или нет «машину» клеточного цикла и начинать деление.
Если сигналы, указывающие клетке, что она должна разделиться, имеют ограниченный характер, клетки Метазоа переходят в неделящееся состояние, которое обозначается как покоящееся, или состояние G0. Из этого состояния многие клетки опять могут войти в цикл при наличии питательных веществ и сигналов, необходимых для наступления деления. Эти сигналы включают соответствующие ростовые факторы и гормоны.
Однако, по мере дифференцировки in vivo или в культуре, клетки часто утрачивают способность пролиферировать и могут перейти в состояние постоянного покоя, которое называется старением. Например, полностью дифференцированные нейроны не делятся, даже в присутствии химических стимуляторов роста.
Некоторые внешние факторы могут стимулировать наступление еще одного состояния клетки, которое называется апоптоз. Хотя апоптоз является нормальным процессом, свойственным развивающимся многоклеточным организмам, его можно индуцировать в культуре клеток с помощью специфических внешних сигналов, например с помощью фактора некроза опухоли-а (TNF-а), который связывается со специфическими рецепорами и запускает цепь биохимических процессов, приводящих клетку к гибели.
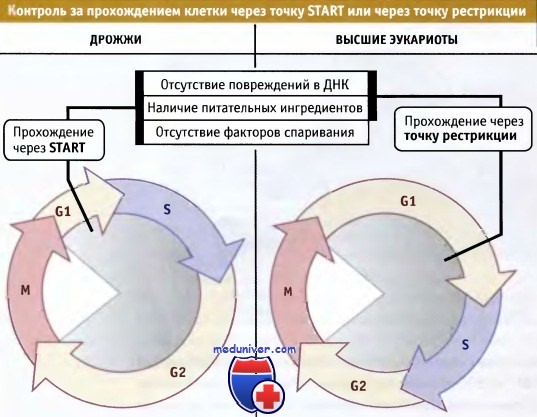
Прохождение клетки через точку START (у дрожжей) или точку рестрикции (у высших эукариот) служит сигналом о готовности клетки войти в цикл и регулируется внешними факторами,
такими как наличие необходимых для роста питательных ингредиентов или отсутствие гормонов спаривания (у дрожжей).
Процесс также регулируется внутренними факторами, например степенью интактности ДНК.
Апоптоз также наступает, когда клетка получает конфликтный внешний сигнал, например сигнал к пролиферации при отсутствии достаточного количества питательных веществ.
У дрожжей, в условиях голодания или в присутствии факторов спаривания, пролиферация подавляется. В ответ на недостаточное питание (недостаток источников азота и углерода) у них снижается активность аденилатциклазы, фермента, превращающего АТФ в цАМФ. Последний представляет собой один из внутриклеточных вторичных мессенджеров, активирующих цАМФ-зависимую про-теинкиназу, которая, в свою очередь, активирует синтез белка и переводит клетку в состояние, коммитированное к делению. Когда в клетках дрожжей уровень цАМФ снижается, они останавливаются в G1-периоде цикла.
Каким образом факторы спаривания ингибируют прохождение клеток по циклу? У гаплоидных почкующихся дрожжей существуют два типа спаривания, МАТа и МАТа, и они секретируют феромоны, на которые реагируют клетки противоположного пола. Например, а фактор вызывает остановку МАТа клеток в G1-периоде Связывание а фактора со своим рецептором на поверхности МАТа клеток активирует протеинкиназный каскад и вызывает протекание двух процессов: экспрессию генов, ответственных за процесс спаривания, и остановку клеточного цикла. Остановка цикла происходит за счет связывания CKI, Far1p с G1 CDK-циклиновыми комплексами.
Если стимулировать покоящиеся клетки к делению, они начинают повторный вход в цикл со стадии G1. До достижения определенной точки в G1-периоде, удаление стимулирующего сигнала предотвращает дальнейшее прохождение клеток по циклу и возвращает их в состояние покоя. Позже, в G1-периоде, клетка проходит определенную точку, после чего удаление сигнала более уже не влияет на ее дальнейшее прохождение по циклу. Таким образом, клетка становится необратимо комми-тированной к циклу деления, независимо от наличия внешних сигналов.
Эта точка в G1-периоде, после которой клетка коммитируется к началу (и завершению) цикла деления, у дрожжей носит название START, а у многоклеточных эукариот точка рестрикции. Схема, иллюстрирующая расположение точек комитирования в клеточном цикле, представлена на рисунке ниже.
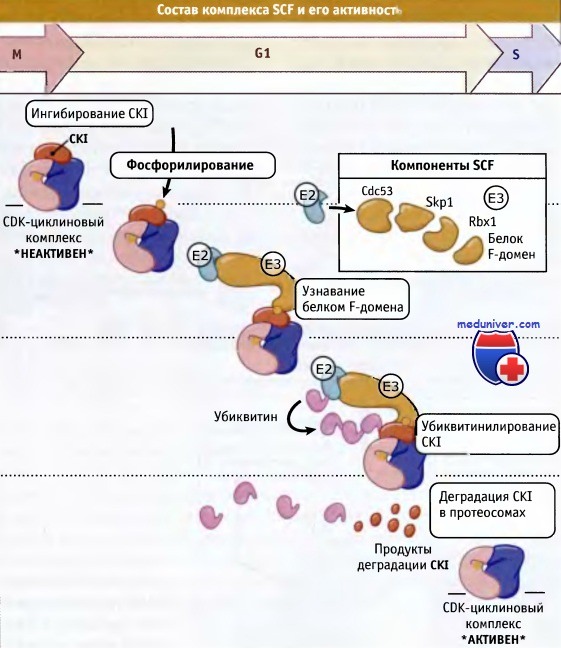
Каким образом происходит реактивация машины, контролирующей клеточный цикл, когда клетка выходит из фазы покоя? Один механизм учитывает снятие ингибирования G1 CDK-циклиновых комплексов. Это достигается, главным образом, за счет деградации ингибиторов CKI, накопленных при остановке клеточного цикла. Деградация осуществляется с участием убиквитина. Убиквитин-лигаза, Е3, необходимая для протеолиза CKI, присутствует во всех клетках и называется SCF. Она отличается от Е3 убиквитин-лигазы, которая участвует в убиквитинилировании митотического циклина.
SCF состоит из четырех коровых субъединиц: Skp1, Cdc53, белка F-домена и белка Rbx 1, содержащего домен пальцев RING. Множественные белки F-домена находятся во всех типах клеток и связываются с различными субстратами, обеспечивая тем самым субстратную специфичность для комплексов SNF. Белок, содержащий домен RING, взаимодействует с ферментом Е2.
Необходимым условием для узнавания субстрата белком F-домена и SCF служит его предварительное фосфорилирование. В случае CKI, фосфорилирование его по нескольким сайтам CDK-циклиновым комплексом позволяет SCF узнать его и деградировать путем протеолиза. Когда CKI деградирует, G1 CDK становится активной. Таким образом, происходит амплификация незначительных количеств активного комплекса CDK-циклина, пока, наконец, активность CDK достигает того уровня, который необходим для индукции перехода через стадию клеточного цикла.
Поэтому некоторые SCF комплексы действуют как негативные регуляторы CKI и проявляют себя как положительные регуляторы, возвращающие клетку в цикл. На рисунке ниже схематически представлен общий состав комплекса SCF и его активность в отношении CKI. Суммируя вышесказаное, отметим, что решение клетки вступать в деление зависит от внешних условий. Если они неблагоприятны для деления, то клетки переходят в состояние покоя. При создании соответствующих условий клетки повторно вступают в G1-фазу цикла. Повторное вступление в цикл и прохождение по фазе G1 требует активации G1 CDK, что и происходит при образовании циклинов в G1-фазе и при инактивации CKI, ингибирующих комплексы CDK-циклин.
Комплекс SCF состоит из четырех основных субъединиц:
Rbx1 (белок, содержащий домен RING), Cullin (например, Cdc53), Skpl и F-домен.
Субъединицы F-домена узнают специфические фосфорилированные субстраты и связывают их с комплексом SCF, также связывая Skp1 через домен F.
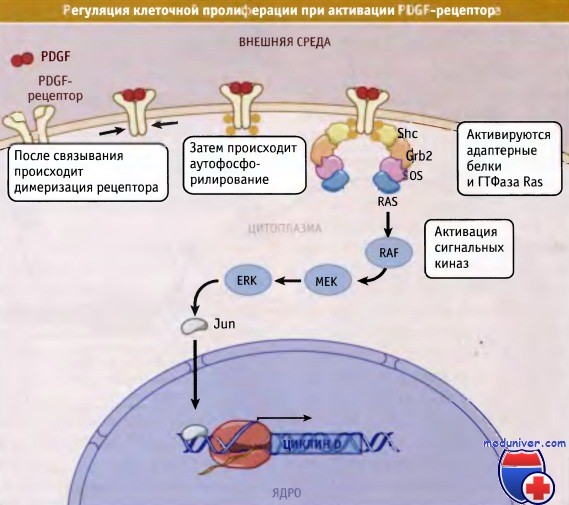
Связывание лиганда с рецептором на наружной мембране клетки приводит к димеризации и к активации рецептора.
В представленном примере рецептор Tyr-киназы, рецептор PDGF, подвергается внутримолекулярному фосфорилированию,
что активирует адаптерные белки Shc, Grb2 и SOS. Эти белки активируют ГТФазу, называемую Ras.
В результате происходит активация сигнального каскада киназ, включающего киназы RAF, MEK и ERK,
которые, в конце концов, активируют транскрипцию.
При этом индуцируется экспрессия генов, регулирующих пролиферацию и прохождение клеток через точку рестрикции, что обеспечивает вступление клетки в цикл деления.