Строение и функции клатриновых везикул в эндоцитозе
• Поэтапная сборка клатриновых трискелионов может представлять собой механический способ преобразования мембран в окаймленные ямки
• Различные адапторные комплексы позволяют отбирать карго для транспорта при связывании как с сигналами сортировки, так и с клатриновыми трискелионами
• ГТФазы из группы динаминов способствуют высвобождению окаймленных везикул из мембраны
• Специальные АТФазы удаляют клатриновое окаймление до начала этапов причаливания и слияния везикул
Эндоцитоз является основным путем, посредством которого в клетку попадают внеклеточные макромолекулы и белки плазматической мембраны. Эндоцитоз и различные его формы представляют собой еще один способ мембранного транспорта, при котором образование транспортных везикул происходит в местах инвагинации плазматической мембраны. По направленности эндоцитоз противоположен общей схеме образования секреторных везикул из мембран ор-ганелл, но в остальном от нее не отличается.
К наиболее изученному механизму эндоцитоза относится протекающий с участием транспортных везикул, покрытых белком клатрином и одним или несколькими адаптерными комплексами, связывающими клатрин с мембраной. Эти адаптеры также участвуют в отборе карго для включения его в образующиеся везикулы, участвующие в эндоцитозе. Клатрин и белки адаптерного комплекса поступают в образующиеся везикулы из цитозоля.
Молекулы карго, находящиеся в клатриновых везикулах, которые образуются на плазматической мембране, часто представляют собой рецепторы внеклеточных лигандов, предназначенные для быстрого проникновения их в клетку. Фактически селективный захват рецепторов клатриновыми везикулами являлся первым примером отбора карго в процессе мембранного транспорта. Клатри-новые везикулы также участвуют в транспортных процессах через транс-Гольджи сеть; эти везикулы имеют другие адаптеры и несут другие белки карго, чем те, которые участвуют в эндоцитозе.
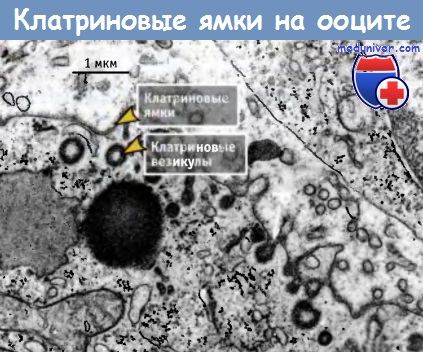
Впервые окаймленные ямки и везикулы были обнаружены при электронно-микроскопическом исследовании ооцитов комаров в начале 1960-х годов. Ооциты захватывают и хранят желточные гранулы, необходимые для дальнейшего эмбриогенеза, и скорость захвата так высока, что большая часть поверхности ооцита содержит ямки, покрытые клатрином.
Фотография клатриновых ямок и везикул,
расположенных на поверхности ооцита, сделанная с помощью трансмиссионного электронного микроскопа.
В начале 1970-х годов была выяснена структура окаймляющего слоя. Это было сделано на синапсах нейронов при рециклизации синаптических пузырьков пресинаптических мембран, используя метод замораживания-замещения. Окаймление выглядит в виде переплетенной сети белковых волокон.
Благодаря такой структуре, окаймление получило название клатрин (лат. clatbratus — защищенный решеткой).
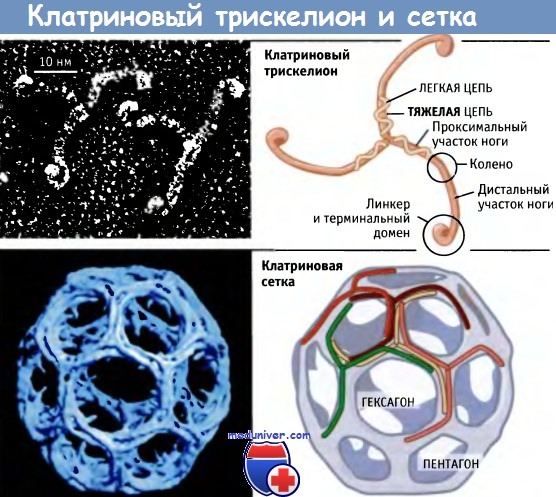
Впервые клатриновые везикулы были выделены из ткани головного мозга в 1969 г., и позже был показано, что клатриновое покрытие состоит в основном из двух белков, тяжелой (180 кДа) и легкой (30 кДа) цепей клатрина, представленных в эквимолярном количестве. Исследования в электронном микроскопе показали, что клатриновые белки образуют трехногую фигуру, называемую трискелион. Каждая «нога» трискелиона включает по одной тяжелой и легкой цепи клатрина. Проксимальный участок ноги расположен ближе к вершине трискелиона и представляет собой место присоединения легких цепей. Дистальный участок располагается дальше от вершины и состоит из тяжелой цепи.
Хотя in vivo клатриновое покрытие мембран может собираться с участием других белков, in vitro клатрин обнаруживает замечательную способность к самосборке в решетчатые структуры. При инкубации выделенных трискелионов в среде с определенным значением pH и солевым составом, они спонтанно образуют пустую клатриновую сетку, которая выглядит так же, как и покрытие, окружающее мембрану in vivo. Сборка клатриновых решеток из очищенных препаратов легких и тяжелых цепей показывает, что трискелион образует гексагональные и пентагональные структуры, представленные на рис. 4.36. Эти структуры образуют сетки, которые напоминают покрытие везикул in vivo.
Таким образом, для образования решетчатой структуры необходимо и достаточно наличие тяжелых и легких цепей клатрина. Каждая вершина представляет собой узел одного трискелиона, а каждая сторона Пентагона или гексагона состоит из четырех ног, принадлежащих различным трискелионам.
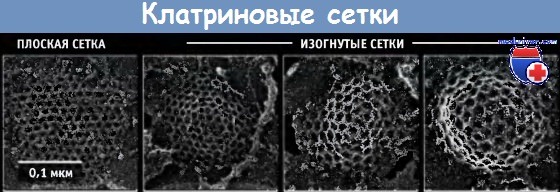
Электронные микрофотографии структур,
окаймленных клатрином, находящихся на плазматической мембране.
Клатриновые сетки имеют различную кривизну.
На каждой фотографии изображена разная сетка.
Геометрически структура выглядит так, что гексагональные клатриновые решетки являются плоскими, но образование пентагонов вызывает изгиб необходимый для формирования сетки. Однако вся последовательность событий, приводящих к образованию полностью изогнутых и окаймленных ямок, in vivo остается неизвестной. Согласно одной из предложенных моделей, часть гексагонов в плоскости каким-то образом меняет структуру и образует пентагоны, что приводит к возникновению кривизны. Другая модель предполагает, что необходимая кривизна мембраны обеспечивается белками, отличными от клатрина, и эта кривизна «фиксируется» образованием пентагонов.
Для сборки клатриновых сеток источники энергии не требуются. Поэтому предполагают, что при сборке происходит изменение свободной энергии, способствующее инвагинации мембраны и образованию везикулы. Однако в деформации плазматической мембраны до образования окаймленной везикулы ключевую роль также играют адаптерные комплексы.
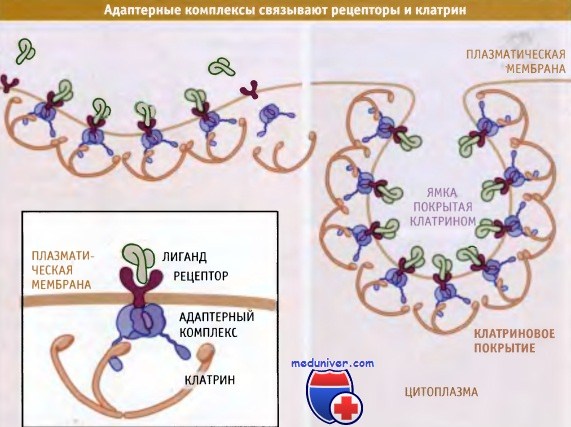
Клатриновое покрытие непосредственно не взаимодействует с мембраной или с мембранными белками. Как показано на рисунке ниже, различные адапторные комплексы являются посредниками между клатриновыми субъединицами и сигналами сортировки, расположенными в цитоплазматическом участке молекул белков карго. На разных этапах транспорта разные адаптеры выбирают трансмембранные белки.
Для завершения эндоцитоза отпочковывающиеся клатриновые везикулы должны отшнуроваться от плазматической мембраны. Процесс отшнуровки требует энергетических затрат и происходит с участием ГТФазы, называемой динамин, которая была впервые обнаружена в клетках мутанта Drosophila shibire. Эти мухи не способны летать при непермиссивной температуре. В клетках мутанта при непермиссивной температуре окаймленные ямки в синапсах нейронов соединены с длинными перемычками и не отшнурованы от плазматической мембраны.
Такие структуры не обнаруживались при пермиссивной температуре. Эти наблюдения позволили предположить, что фентотип shibire-мутанта обусловлен неспособностью компонентов синаптического пузырька передаваться через плазматическую мембрану, тем самым блокируя передачу нервного импульса. Мутация shibire вызвана дефектом динамина. В присутствии негидролизуемого ГТФ аналога очищенный динамин клеток млекопитающих in vitro образует на плазматической мембране эти длинные перемычки.
Так называемые «воротнички» представляют собой по сравнению с shibire небольшие по размеру структуры, состоящие из динамина, которые располагаются вокруг перемычек нормальных клатриновых ямок. Динамин поступает из цитоплазмы, группируясь вокруг перемычки отпочковывающейся окаймленной везикулы в виде спирали с тем, чтобы помочь отшнуровке клатриновых ямок от плазматической мембраны.
Однако механизм отшнуровки неизвестен. Согласно одному представлению, при гидролизе ГТФ эти спиральные структуры сжимаются, тем самым обеспечивая слияние мембраны, необходимое для высвобождения новых клатриновых пузырьков. Другая модель предполагает, что динамин косвенным образом способствует отшнуровке, по-видимому, привлекая другие молекулы или изменяя конформацию окаймленной ямки в сторону образования везикулы. Какова бы ни была причина, очевидно, что динамин является основным компонентом процесса высвобождения клатриновых везикул из плазматических мембран, и вероятно, также из транс-Гольджи сети.
После отделения от мембраны клатриновые везикулы взаимодействуют с элементами актинового цитоскелета, что способствует продвижению везикул по цитозолю. Вновь образованные везикулы могут служить местами сборки актиновых филаментов, обеспечивающих это продвижение. Хотя еще остается выяснить, насколько такое движение необходимо, очевидно, что многие «дополнительные белки», которые участвуют в образовании клатриновых ямок, в частности динамин, могут нуклеи-ровать сборку актина.
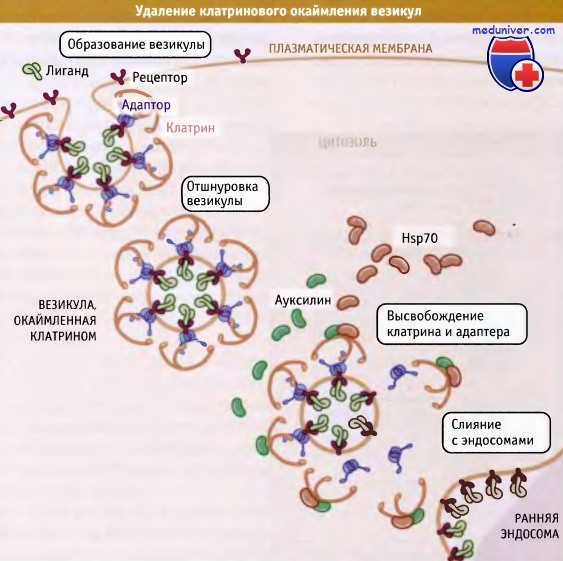
После образования клатриновой везикулы окаймление удаляется с тем, чтобы обеспечить везикуле возможность позиционироваться на следующем компарт-менте, ранней эндосоме. В удалении клатрина участвуют по крайней мере два белка, «раздевающая АТФаза», которая является цитозольным ферментом из группы белков теплового шока, Hsp70, и ауксилин, белок, взаимодействующий с этой АТФазой. Точный механизм удаления окаймления неизвестен, однако, вероятно, он включает в себя дестабилизацию и отделение клатринового слоя после присоединения Hsp70.
Такой механизм согласуется со способностью этих АТФаз, участвующих в качестве шаперонов при синтезе и деградации молекул белка, связываться со структурами высшего порядка и вызывать их дестабилизацию. Определенную роль в отделении окаймления, особенно при высвобождении адапторных комплексов, может играть гидролиз кислых фосфолипидов.
Кпатриновые трискелионы собраны в замкнутые корзиноподобные структуры.
Эти структуры могут образовывать сеть, состоящую из многоугольников пентагонального и гексагонального типа,
и, таким образом, генерируются замкнутые корзиноподобные структуры.
Для простоты на нижнем правом рисунке легкие цепи не показаны.
Клатрин связывается с адаптерами,
которые узнают сигнал сортировки, расположенный на цитоплазматическом участке предназначенных для транспорта белков.
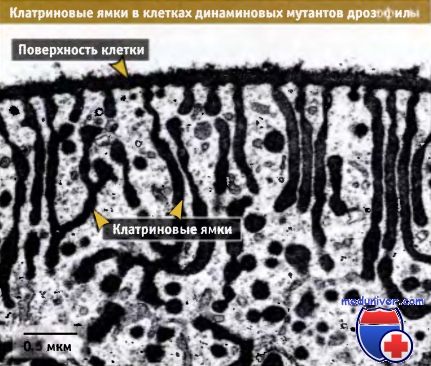
Электронная микрофотография клатриновых ямок в клетках мутанта дрозофилы, дефектных по динамину.
Мутантные мухи не были способны летать при непермиссивной температуре 30 °С.
Видны канальцы, распространяющиеся от клеточной поверхности и заканчивающиеся клатрин-окаймленными утолщениями.
Эти утолщения не могут отшнуровываться, поскольку динаминовые мутанты не способны нормально функционировать при непермиссивной температуре.
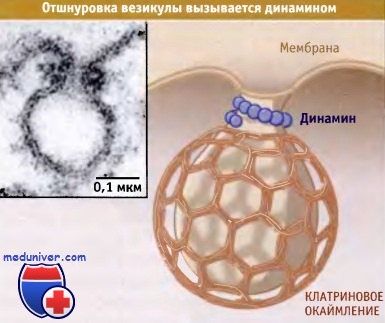
Динамин может образовывать спиральные структуры вокруг перетяжки (шейки)
отпочковывающейся везикулы, которые вызывают ее отшнуровку.
В удалении клатринового окаймления везикул участвуют два белка: Hsp70 и ауксилин.