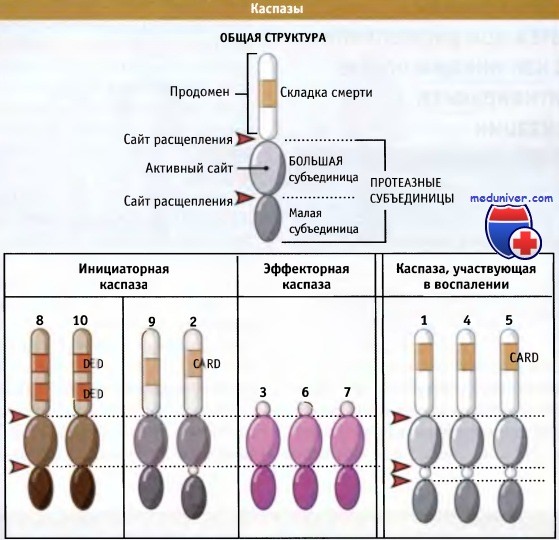
• Протеазы, называемые «каспазами», подразделяются на три группы: инициаторные, эффекторные и участвующие в воспалении. Два первых типа каспаз связаны с апоптозом
• Морфологические и биохимические особенности апоптотических клеток обусловливаются действием эффекторных каспаз на соответствующие субстраты
• Обнаружено много субстратов каспаз, и в некоторых случаях известно, какие клеточные эффекты вызываны их расщеплением
Каспазы — цистеин содержащие специфические протеазы, атакующие (расщепляющие) субстраты после ас-партата, регулируют многие клеточные и биохимические изменения в погибающих апоптотических клетках. В большинстве клеток млекопитающих каспазы находятся в виде неактивных зимогенов; и при активации апоптоза нового их синтеза не происходит.
Существует три основных типа каспаз, обладающих в клетке различными функциями: эффекторные, инициаторные и участвующие в воспалении. Эффекторные каспазы (в основном, каспазы-3 и -7 позвоночных) ответственны за расщепление многих различных белков, чтобы осуществить апоптоз. Обычно эти каспазы расщепляют субстраты, имеющие в своей структуре следующие последовательности аминокислот: Asp-Xaa-Xaa-Asp/Gly, Asp-Xaa-Xaa-Asp/Ser или Asp-Xaa-Xaa-Asp/А1а.(«/» обозначает место разрыва, а Хаа — любую аминокислоту).
Установлено, что в клетках млекопитающих содержится около 500 субстратов каспаз, хотя последствия расщепления большинства из них неизвестны. В некоторых случаях расщепление субстрата играет определенную роль в апоптозе и вызывает изменения, связанные с этой формой клеточной гибели. В других случаях расщепление субстрата не имеет отношения к апоптозу и играет «сопутствующую» роль.
Наблюдаемая при апоптозе фрагментация ДНК является результатом расщепления одного из субстратов каспаз. Одна из клеточных ДНКаз (CAD = orspase-dependent DNase) находится в клетке в виде комплекса с ингибитором, iCAD. Фактически, при действии ингибитора CAD свернута в комплекс, и ее активность проявляется только, когда iCAD расщепляется эффекторной каспазой.
Активная CAD начинает расщеплять ДНК по наиболее доступным местам между нуклеосомами, что и приводит к характерной ее деградации, в виде «лесенки», наблюдаемой при апоптозе. Клетки, у которых отсутствует либо CAD, либо iCAD, при развитии апоптоза не обнаруживают такого характера расщепления ДНК.
Субстратом для эффекторных каспаз служит киназа легкой цепи миозина, ROCK-1. Однако в этом случае при расщеплении фермент активируется. Сходным образом под действием эффекторных каспаз расщепляется и активируется другой фермент, гельзолин. Оба фермента вызывают изменения цитоскелета, в результате чего в клетке развивается «пузырение», характерное для апоптоза.
Хотя каждая реакция расщепления субстратов, катализируемая каспазой, может обусловливать характерные признаки апоптоза, не показано, что какой-либо из субстратов или даже их группа непосредственно участвует в клеточной гибели. В конце концов, каспазы могут вызывать апоптоз в результате расщепления многих различных субстратов, и в настоящее время клеточную гибель вследствие активации каспаз невозможно эффективно блокировать, воздействуя на любой из известных наборов специфических субстратов.
Субстратов, вызывающих гибель, может быть много (что кажется вероятным), или мы просто не выяснили, какие еще являются наиболее важными. В то же время фармакологические ингибиторы каспаз, например валин-аланин-аспартат-фторметилкетон (VAD-fmk) блокирует клеточные и биохимические проявления апоптоза, и в некоторых случаях даже предотвращает гибель клеток.
Схема строения различных типов каспаз клеток позвоночных.
Обратите внимание на существование продоменов и областей белок-белковых взаимодействий у инициаторных каспаз и участвующих в воспалительных процессах.
Эффекторный домен клеточной гибели (DED); домен мобилизации каспаз (CARD).