• Белки проходят через водный канал, состоящий из комплекса Sec61, расположенного в мембране ЭПР
• С каналом связано много дополнительных белков, участвующих в транслокации, формировании нативной структуры и в различных модификациях
Белки преодолевают мембрану ЭПР по заполненному водой каналу, который проходит через мембрану и специально предназначен для транслокации. Этот канал, вместе со связанными с ним другими белками, называется транслоконом. Транслоконовый комплекс обладает динамичной структурой, и прежде чем рассмотреть транслокацию, целесообразно подробнее ознакомиться с его строением.
Оказалось крайне трудным доказать присутствие каналов, обеспечивающих транспорт белков через мембрану ЭПР. Основные положения сигнальной гипотезы предполагали существование каналов, однако в то время высказывались также и другие представления, например о непосредственном транспорте секреторных белков через липидный бислой. Первое убедительное доказательство существования каналов было получено на основании электрофизиологических экспериментов.
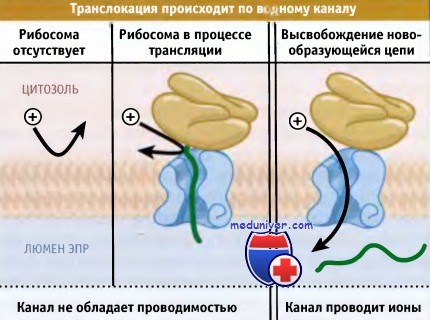
В них исследовали способность ионов транспортироваться через мембраны везикул (называемых микросомы), образованных из гранулярного ЭПР. Ионы не проходят через липидный бислой, и поэтому если регистрируется проводимость, то это свидетельствует о присутствии в мембране каналов. Микросомы, в которых не наблюдалась транслокация (из-за удаления связанных с ними рибосом), обнаруживали лишь незначительную электропроводимость. Аналогичным образом, ионы не могли пройти через мембрану, занятую рибосомами, находящимися в процессе транслокации.
Ионы проходят через мембрану ЭПР, только когда цепи переносимых белков освобождаются от связанных рибосом.
Это позволяет предполагать, что новообразующаяся цепь проходит через мембрану по водному каналу.
Для того чтобы канал оставался открытым, необходимо присутствие рибосомы, поскольку,
если после высвобождения новообразующейся цепи рибосома удаляется, канал теряет проводимость.
Однако, как иллюстрирует рисунок ниже, можно было зарегистрировать проводимость, если насцентные белки высвобождались из рибосом, которые оставались связанными с мембраной.
Этот эксперимент позволил прийти к следующим двум выводам:
• Ионы способны проходить через водные каналы, которые стабилизируются рибосомами, а при отсутствии рибосом ионы не проходят.
• Новообразованная цепь белка занимает этот канал; причем ионы могут пройти через канал только при высвобождении новообразованной цепи.
В последующих экспериментах с использованием других методов было подтверждено существование канала, т. е. прямо показано, что новообразованные секреторные белки пересекают мембрану, находясь в водном окружении.
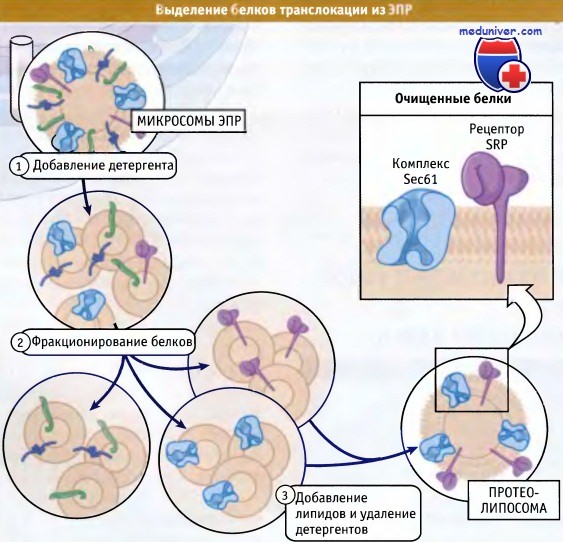
Какие белки входят в состав канала транслокации? Кандидаты должны удовлетворять нескольким критериям, включая близость к транслоцируемому белку и взаимодействие с мембрано-связанными рибосомами. Для того чтобы обнаружить белки, удовлетворяющие этим критериям, микросомы обрабатывали детергентом, т. е. переводили мембранные белки в растворимую форму. На рисунке ниже показано дальнейшее фракционирование белков с целью найти минимальный набор компонентов, необходимых для транслокации.
Индивидуальные белки заключали в липидные везикулы, называемые протеолипосомы, которые в экспериментах по транслокации in vitro моделировали микросомы. Таким образом, можно было контролировать белковый состав этого искусственного ЭПР и выяснить роль каждого белка в транслокации. Концептуально такой подход напоминает разборку и повторную сборку автомобиля с тем, чтобы найти только такие сочетания деталей, которые необходимы для того, чтобы автомобиль мог передвигаться. Так, двигатель и некоторые другие детали абсолютно необходимы, а такие регуляторные приспособления, как тормоз, могут отсутствовать.
Этот подход продемонстрировал, что для транслокации некоторых белков необходимы только SRP, SR и комплекс трех трансмембранных белков, обозначаемый Sec61. При этом уже было известно, что SRP и SR участвуют в адресовании, и комплекс Sec61 представляет собой подходящего кандидата на роль каналообразующего белка, через который осуществляется транслокация.
Sec6l обладает консервативной структурой, что свидетельствует о его критической роли в процессе транслокации. Впервые, белки, соответствующие этому комплексу, были обнаружены в дрожжах, при генетическом скрининге, специально предпринятом для поиска генов, кодирующих компоненты, необходимые для входа секреторных белков в ЭПР. Было обнаружено несколько таких генов, включая SEC61. Этот ген кодирует белок Sec61p, представляющий собой интегральный белок мембраны, который десять раз пронизывает мембрану ЭПР. В клетках млекопитающих присутствует гомолог этого белка, Sec61a.
Для диссоциации микросом ЭПР на небольшие мицеллы белков и липидов можно использовать детергенты.
В дальнейшем мицеллы можно подвергнуть фракционированию для выделения необходимых белков.
После удаления детергента и добавления новых липидов образуются везикулы, которые содержат только необходимые белки.
На основании биохимических экспериментов in vitro предположили, что Sec61p окружает транслоцируемый белок и очень вероятно, что он образует стенку канала. Генетические и биохимические исследования показали, что Sec61p прочно связывается с двумя другими, более мелкими белками, точные функции которых неизвестны:
• Sss1p (Sec61γ у млекопитающих) и
• Sbh1р (Sec61b у млекопитающих)
Вместе три этих компонента образуют гетеротримерный комплекс Sec61.
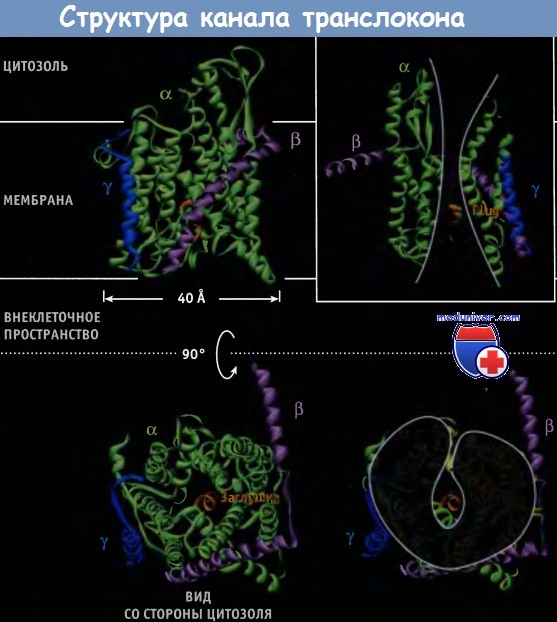
Детальное строение комплекса Sec61 было установлено с использованием данных, полученных для SecY комплекса архебактерий Methanococcus jannascbii, структурно и функционально близкого к Sec61 эукариот. Особенности строения комплекса позволяют выделить несколько важных характеристик канала транслокации. Прежде всего, необходимо отметить, что внутренняя часть канала состоит из трансмембранных доменов Sec61 и по форме напоминает часовое стекло. Отверстие, напоминающее пору, закрыто небольшой заглушкой, которая представляет собой часть структуры Sec61a. Со стороны рибосомы (т. е. цитозоля) трансмембранные домены 1-5 Sec61a образуют половину структуры, по форме напоминающей раковину моллюска, а трансмембранные домены 6-10 образуют другую половину. Между двумя половинами расположен белок Sec61γ.
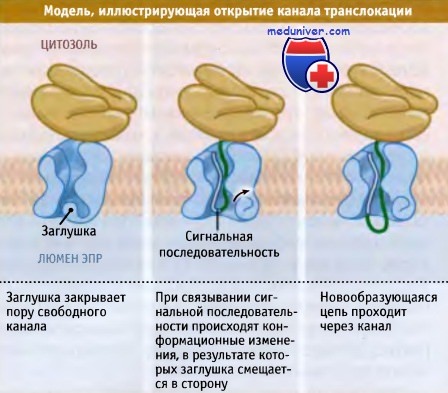
Белковая цепь должна пройти через центральную пору канала; для этого отверстие должно быть открыто. Для выяснения механизма открытия поры были проведены биохимические эксперименты, позволившие выявить участки канала, примыкающие друг к другу. Первое событие начала транслокации заключается в узнавании сигнальной последовательности канальными белками. Вероятно, при этом заглушка канала удаляется. Важно отметить, что узнавание происходит только после того, как белковая цепь уже заняла канал, т.е. когда в поре находится белок, а не заглушка. Отметим, что пора канала довольно узкая и окружена гибкими остатками гидрофобных аминокислот. Поэтому проходящая через пору белковая цепь, вероятно, занимает все пространство, и транспорт ионов через канал сильно затруднен.
Еще до выяснения строения комплекса Sec61 определили, что он представляет собой тример или тетрамер, и предполагали, что для формирования канала необходимо несколько копий комплекса. Однако в настоящее время это представляется маловероятным, и скорее всего, канал образует одна копия Sec61 (т. е. по одной молекуле Sec61a, Sec61b и Sec61γ). Причина, по которой для формирования канала необходимо участие трех или четырех копий Sec61, пока остается неясной.
Хотя Sec61 непосредственно образует канал, рядом находятся другие белки, и комплекс Sec6l может рассматриваться как некий каркас, на котором собираются белки ЭПР, принимающие участие в переносе, а также в модификации и формировании нативной структуры белка. Например, в каждом активном транслоконе, наряду с рибосомой и описанным выше SR комплексом, присутствует сигнальная пептидаза, которая отщепляет сигнальную последовательность от переносимого белка. Также в транслоконе находится олигосахарид — трансфераза (ОСТ), представляющая собой комплекс ферментов, которые ковалентной связью присоединяют сахарные остатки к белковым цепям. С растущей цепью взаимодействуют и другие белки, функции которых остаются невыясненными.
С сигнальной последовательностью и трансмембранными доменами часто связан т. н. «мембранный белок, ассоциированный с переносимым белком (TRAM)». В отличие от ферментов, деградирующих или модифицирующих участки образующейся цепи, белок TRAM необходим для переноса некоторых белков на нескольких этапах этого процесса. Еще один белковый комплекс, принимающий участие в переносе, называется белок, ассоциированный с транслоконом (TRAP). Он также распространен, как Sec61a, и облегчает узнавание сигнальной последовательности канальными белками, хотя механизм его действия остается неизвестным. Очевидно, что транспортный канал не функционирует изолированно, а, по-видимому, является частью большого белкового комплекса, который регулирует процесс транслокации. Таким образом, при необходимости, клетка в любой момент времени может изменить процесс транслокации индивидуальных белков.
Стенки канала по форме напоминают часовые стекла.
Центральное отверстие поры закрыто небольшой заглушкой, которая открывается после связывания сигнальной последовательности с белками канала.
В рамке справа удалены некоторые трансмембранные спирали и видна пора, стенки которой напоминают по форме часовые стекла.
Со стороны цитозоля видны трансмембранные спирали, расположенные в виде раковины.
Связывание сигнальной последовательности с канальными белками, вероятно,
вызывает конформационные изменения, которые приводят к открытию канала и позволяют белковой цепи проходить через него.