• К+-каналы функционируют как заполненные водой поры и катализируют селективный и быстрый транспорт ионов К+
• К+-канал представляет собой комплекс, состоящий из четырех идентичных субъединиц, каждая из которых участвует в образовании поры
• Селективный фильтр К+-канала представляет собой эволюционно-консервативную структуру
• Селективный фильтр К+-канала катализирует процесс дегидратации ионов, благодаря чему канал становится специфичным и транспорт ионов по нему значительно ускоряется
Калиевые (К+) каналы представляют собой белки, интегрированные в плазматическую мембрану, которые обеспечивают выход ионов К+ из клетки. Калиевые каналы — эволюционно-консервативные структуры в клетках всех живых существ, от бактерий до человека.
В различных типах клеток они выполняют разнообразные функции, от поддержания мембранного потенциала покоя до завершения потенциала действия и поддержания баланса электролитов. (У прокариот функции К+-каналов еще недостаточно выяснены.)
Подобно другим ионным каналам, К+-каналы имеют разные воротные механизмы, т. е. в ответ на действие электрических или химических сигналов они могут открываться и закрываться. Поскольку клетки поддерживают существенно более высокую концентрацию внутриклеточного калия, открытие калиевых каналов приводит к выходу его из клетки, так как ионы движутся в направлении электрохимического градиента.
Так же как и другие ионные каналы, калиевые представляют собой узкие заполненные водой поры, через которые происходит транспорт ионов. К+-каналы проявляют высокую избирательность по отношению к ионам калия. Эта избирательность играет важную роль в поддержании постоянства концентрации внутриклеточных ионов.
К+-каналы представляют собой тетрамеры, состоящие из идентичных субъединиц,
каждая из которых участвует в образовании центральной поры.
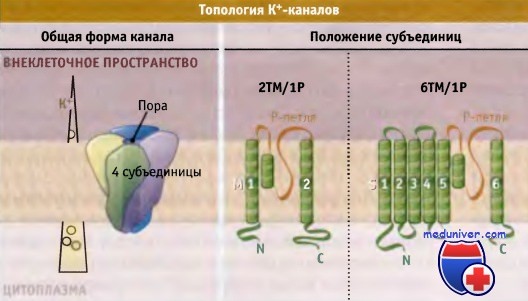
Существуют два основных типа субъединиц, состоящие из двух и из шести трансмембранных доменов.
Оба типа имеют поровую петлю. Слева показано направление градиента К+ в покоящихся клетках млекопитающих.
По данным электрофизиологических исследований, транспорт К+ через каналы происходит со скоростью порядка 108 ионов в 1 с, что близко к максимальной скорости диффузии в воде молекул растворенного вещества. В настоящем разделе мы рассмотрим молекулярные основы селективного и при этом быстрого транспорта ионов К+ через мембрану клетки.
Каждый К+-канал представляет собой гомотетрамер, состоящий из четырех субъединиц, образующих центральную пору, через которую осуществляется транспорт ионов. В зависимости от трансмембранной топологии субъединиц различают два основных типа К+-каналов. Тип 2ТМ/1Р характеризуется тем, что каждая субъединица имеет две трансмембранных (2ТМ) а-спирали, M1 и М2, которые разделены петлей поры (Р).
Р-петля содержит короткий спиральный фрагмент, который называется спиралью поры. Эти детали строения являются общими для всех К+-каналов. Тип 2ТМ/1Р включает канал внутреннего выпрямления, КАТР, и G-белок, с которым сопряжен К+-канал. Каждая субъединица еще одного типа К+-канала, который обозначается как 6ТМ/1Р, содержит шесть трансмембранных доменов (6ТМ) и одну Р-петлю.
В этом типе К+-канала домены S5-P-S6 образуют область поры таким же образом, как М1-Р-М2 домены в каналах типа 2ТМ/Р. Четыре добавочных трансмембранных сегмента (S1-S4) участвуют в воротном механизме. Каналы типа 6ТМ/1Р включают К+-каналы, зависящие от потенциала на мембране (Kv-каналы) и от лиганда, например Са2+ активируемые К+-каналы.
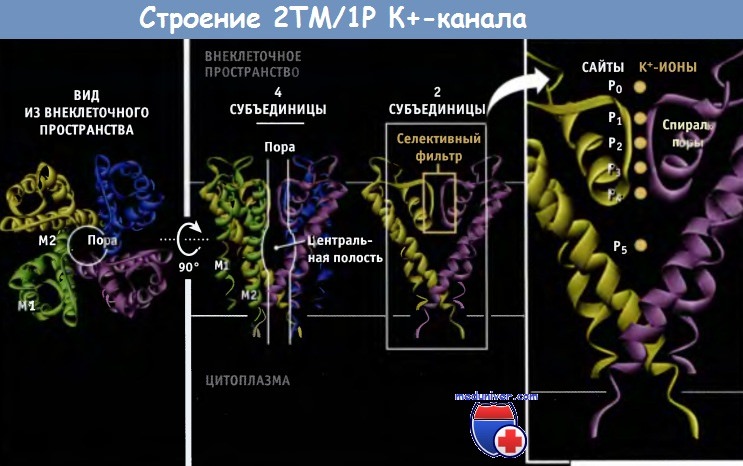
Нам удалось выяснить причины высокой селективности К+-каналов на основании электрофизиологических исследований, а также используя мутагенез и рентгеноструктурный анализ. Данные рентгеноструктурного анализа позволили не только подтвердить ранее высказанные предположения, но дали возможность создать модели каналов на атомарном уровне. Первый К+-канал, для которого была установлена кристаллическая структура, представлял собой KcsA канал бактерий, относящийся к 2ТМ/1Р типу, в закрытой конфигурации.
Пора, через которую проходят ионы, состоит из двух основных частей, селективного фильтра и центральной полости. Внутри поры расположены М2 (внутренние) спирали и Р-петли. Селективный фильтр, который представляет собой самую узкую часть поры, расположен ближе к ее внешнему отверстию и связывает ионы К+. Длина его составляет 12 А, а ширина 3 А, и каждая субъединица добавляет одну Р-петлю к его структуре.
В канале присутствуют шесть ион-содержащих сайтов: четыре из них (Р1-Р4) расположены в пределах селективного фильтра, а из двух остальных (Р0 и Р5) один расположен на внешней стороне фильтра, и один — в центральной полости. В составе кристаллической структуры эти сайты формируют сложное изображение всех белков К+ канала.
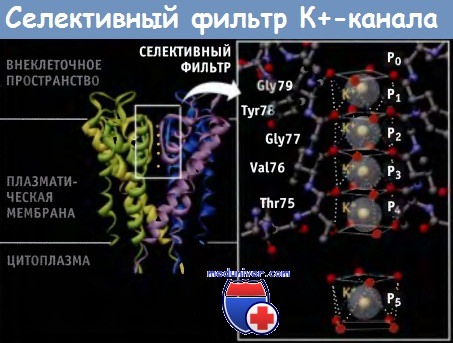
Каждая из четырех Р-петель селективного фильтра К+ канала содержит высококонсервативную последовательность, которая называется сигнатурной последовательностью: Thr-X-Gly-Tyr-Gly или Thr-X-Gly-Phe-Gly (TXGYG или TXGFG соответственно), где X представляет собой один из нескольких аминокислотных остатков. Сигнатурная последовательность для канала KcsA имеет следующий вид: Thr-Val-Gly-Tyr-Gly (TVGYG).
От каждого из аминокислотных остатков в канал попадают атомы карбонильного кислорода (или из основной направлены молекулы, или из боковой цепи). Как показано на рисунке ниже, эти атомы, которые частично несут отрицательный заряд, располагаются по углам четырех коробчатых структур, которые координируют положение проходящих ионов К+. Скоординированное положение ионов К+ в селективном фильтре напоминает линейную цепь.
Ленточная модель кристаллической структуры KcsA К+-канала, состоящей из белков.
Отмечены предполагаемые границы мембраны. Для простоты на двух рисунках справа изображены только две субъединицы.
Модели построены на основании данных Protein Data Bank file 1K4C.
Представлена структура укороченного KcsA-канала и некоторые N- и С-терминальные участки не показаны.
Ионы К+, находящиеся в растворе, гидратированы, т. е. окружены молекулами воды. Для того чтобы ионы могли проникнуть в узкий селективный фильтр, они должны от этих молекул освободиться. Однако процесс дегидратации ионов требует значительного количества энергии, поскольку при этом необходимо разорвать силы притяжения между положительно заряженными ионами К+ и частичным отрицательным зарядом дипольных молекул воды. Эта проблема решается с помощью К+-канала.
Отрицательный заряд атомов кислорода, находящихся в селективном фильтре, действует таким же образом, как молекулы воды, создавая гидрофильное окружение, которое снижает энергию дегидратации для ионов К+ и копирует слабые отрицательные заряды диполей воды, создающих гидратную оболочку.
Дегидратация является основным этапом на пути формирования ионной селективности каналов. Рассмотрим другой распространенный одновалентный катион, Na+, который по размеру меньше иона К+, однако в 104 раз медленнее проникает через К+-каналы. Причина того, что К+-каналы практически непроницаемы для ионов Na+, заключается в том, что размер селективного фильтра калиевого канала компенсирует энергию дегидратации ионов К+, но не ионов натрия.
Дегидратированные ионы К+ хорошо проходят через селективный фильтр, однако дегидратированные ионы Na+ оказываются слишком мелкими для того, чтобы взаимодействовать с карбонильным кислородом таким образом, чтобы дегидратация оказалась энергетически выгодной. Поэтому структура селективного фильтра способствует прохождению ионов калия, но не ионов натрия.
Для того чтобы скорость диффузии ионов через К+-канал составляла 108 ионов в 1 с, процессы их дегидратации, прохождения через селективный фильтр и гидратации должны происходить за 10 нс. С помощью компьютерного моделирования разработана модель, позволяющая рассчитать максимально возможную скорость движения ионов. Модель была разработана с учетом анализа различных кристаллических структур, причем каждая структура соответствовала определенной концентрации ионов К+.
Все четыре субъединицы KcsA К+-канала бактерий участвуют в формировании селективного фильтра.
Для простоты показаны остатки поровой петли только двух субъединиц вместе с соответствующими атомами кислорода поровых петель двух других субъединиц.
Р0-Р5 представляют собой шесть предполагаемых сайтов связывания ионов К+. Р0 сайт расположен на наружном устье поры канала (ион К+ не показан).
Каждый из Р1-Р4 сайтов селективного фильтра образован восемью атомами кислорода (помечены красным цветом).
Р-петель. Сайт Р5 в центральной полости занят гидратированным ионом К+ (атомы кислорода восьми молекул воды помечены красным цветом).
Модели построены на основании данных Protein Data Bank file 1K4C.
Структура учитывает присутствие в кристалле всех белков, поэтому все сайты заняты ионами К+.
Показано предполагаемое положение мембраны.
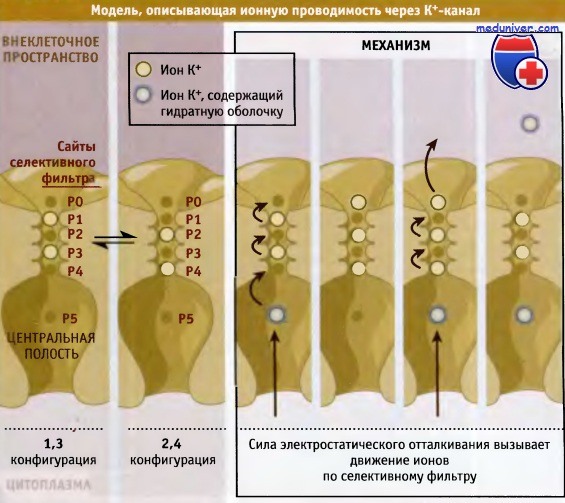
При разработке модели исходили из предположения, что в каждый данный момент времени ионы калия в селективном фильтре могут занимать или положения Р1 и Р3 (конфигурация 1,3), или Р2 и Р4 (конфигурация 2,4). Эти конфигурации предполагают, что ионы К+ не располагаются близко друг к другу из-за существования сил взаимного отталкивания. Две конфигурации энергетически близки друг к другу и находятся во взаимном равновесии.
Как только ионы К+ из центральной полости поступают в селективный фильтр, силы электростатического отталкивания продвигают их по фильтру по пути одной из двух возможных конфигураций. В результате ионы выходят с внешней стороны канала. Высокая проводимость канала обеспечивается в том числе отсутствием энергетического барьера между конфигурациями 1,3 и 2,4, который при его наличии существенно затруднял бы транспорт.
Отметим также, что общий принцип, определяющий быстрый транспорт ионов, заключается в том, что при открытии канала поры расширяются и через них свободно проходят ионы, которые достигают селективного фильтра. Так, для К+-каналов эффективное расстояние, на которое проникают ионы, укорачивается до 12 А, т. е. до длины селективного фильтра, а не до толщины мембраны.
Другая особенность структуры К+-каналов заключается в существовании центральной полости диаметром 10 А, расположенной с внутриклеточной стороны селективного фильтра. Изнутри полость выстлана гидрофобными остатками, что сводит к минимуму взаимодействие ее стенок с гидратированными ионами. В то же время центральная полость помогает ионам К+ преодолевать силы отталкивания от гидрофобного центра липидного бислоя.
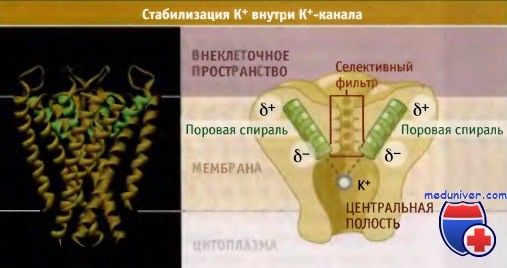
Два характерных свойства центральной полости способствуют стабилизации ионов К+ в канале, в центре мембраны, где существует максимальный энергетический барьер для прохождения ионов. Во-первых, ионы К+ находятся в центральной полости в гидратированном состоянии. Во-вторых, как показано на рисунке ниже, частичный отрицательный заряд поровых спиралей каждой субъединицы канала направлен к центру центральной полости.
(Частичные отрицательный и положительный заряды на а-спиралях возникают в результате образования водородных связей между соседними витками. При этом С-концевая часть приобретает более отрицательный заряд, чем N-концевая.)
Ионы К+ движутся через мембрану в направлении концентрационного градиента, из внутриклеточной среды во внеклеточное пространство.
Модель постулирует, что в каждый данный момент времени в селективном фильтре находятся два частично дегидратированных иона К+.
Эти ионы быстро перемещаются между двумя конфигурациями, которые обладают одинаковой свободной энергией.
Увеличение количества ионов вызывает рост силы электростатического отталкивания, за счет которой ионы продвигаются по селективному фильтру.
Таким образом, селективный фильтр снимает энергетические барьеры и обеспечивает высокую скорость прохождения ионов К+.
Четыре поровых а-спирали (помечены зеленым цветом) примыкают к центральной полости
своими отрицательно заряженными участками диполей и стабилизируют положительно заряженный ион К+,
занимающий сайт Р5 (для простоты на правом рисунке представлены только две поровые спирали).
Показан частичный заряд спиральных диполей.
Модели построены на основании данных Protein Data Bank file 1K4C.