Возможность наблюдать ионные каналы при атомном разрешении ознаменовала собой крупнейший прорыв в исследовании процессов мембранного транспорта. Например, данные рентгеноструктурного анализа совершили переворот в нашем понимании основных принципов селективности и проницаемости для новых типов ионных и аквапориновых каналов. Однако данные рентгеноструктурного анализа получены для относительно небольшого количества мембранных транспортных белков, и во многих случаях мы располагаем результатами, полученными для бактериальных белков, которые можно выделить в достаточных количествах.
Анализ кристаллической структуры других транспортных белков позволяет выяснить более общие принципы их функционирования. Наряду с этим, анализ кристаллической структуры транспортных белков эукариот дает возможность проникнуть вглубь более сложных процессов, таких, например, как функционирование нейронов.
Установление кристаллической структуры оказалось необходимой предпосылкой для разработки моделей, описывающих работу мембранных транспортных белков. Возможность наблюдать за транспортными белками в реальном времени, используя методы ядерного магнитного резонанса (ЯМР) или другие спектроскопические методы, дополняет данные кристаллической структуры. С помощью этих методов можно получить информацию о переходе белков из одной конформации в другую, о регуляции воротного механизма ионного канала и о конформационных изменениях, происходящих с белками-носителями в процессе переноса ими различных метаболитов.
На генетическом и функциональном уровнях обнаружено множество мембранных переносчиков, ионных каналов и насосов. Современные методы молекулярной генетики позволяют идентифицировать все новые транспортные белки. Более того, выяснилось, что наряду с порообразующими субъединицами для функционирования некоторых транспортных белков необходима субъединичная структура. Для более полного понимания регуляции активности транспортных белков была бы важна идентификация других подобных субъединиц.
Микроаррей-технология и протеомный анализ также позволяют сравнить уровень экспрессии транспортных белков и их регуляторов в нормальных клетках и при различных патологиях.
В качестве перспективной задачи представляется крайне необходимым связать информацию, полученную на основании данных анализа структуры, с физиологическими свойствами клеток и организмов. Нам необходимо изучать транспортные белки в окружении клеточных структур и липидов и исследовать механизмы транспорта и его регуляции. Многие заболевания связаны с мутациями в генах, кодирующих синтез мембранных транспортных белков. Однако для понимания роли определенного ионного канала в развитии заболевания важно оценить функцию этого канала в масштабе всей клетки и, наконец, на уровне органа и организма в целом.
Известно, например, что мутации в белке хлорных каналов — регуляторе трансмембранного переноса при кистозном фиброзе (CFTR) являются причиной аномальной реабсорбции Na+ в легких и желудочно-кишечном тракте больных. Это приводит к образованию густой слизи, которая накапливается в органах, зависимых от такого типа транспортной функции клеток эпителия. Подобные исследования требуют разработки новых биологических тестов, позволяющих оценивать специфические функции. Понимание молекулярных механизмов работы различных ионных каналов дает возможность понять детали функционирования клеток нервной системы и выяснить причины развития аномалий, приводящих к аритмиям и внезапной смерти.
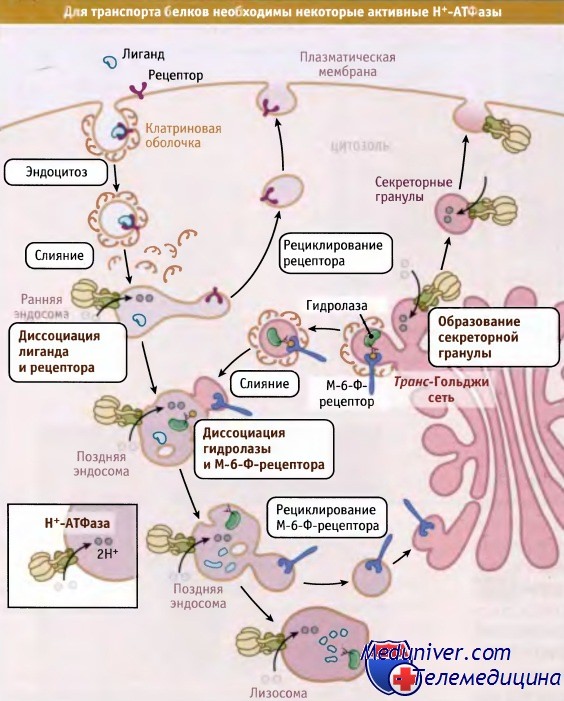
Некоторые V-АТФазы участвуют в везикулярном транспорте.
Закисление среды в эндосомальных компартментах необходимо для диссоциации комплексов белок-рецептор и рециклирования рецепторов.
Наряду с этим, Н+-АТФазы необходимы для образования секреторных гранул.
Были рассмотрены три основных класса мембранных транспортных белков: каналы, переносчики и насосы. Эти белки расположены в плазматической мембране и в мембранах внутриклеточных органелл, таких как эндоплазматический ретикулум, эндосомы, лизосомы и митохондрии. Они принимают участие в многочисленных клеточных процессах, начиная с усвоения питательных компонентов, например глюкозы, и заканчивая более сложными физиологическими задачами, такими как реабсорбция метаболитов клетками почек и распространение потенциала действия.
Каналы обладают порами, которые обеспечивают быструю пассивную диффузию ионов или других метаболитов через биологические мембраны. Они различаются по селективности: некоторые каналы высокоселективны по отношению к К+, Na+, Са2+, Cl- или к молекулам воды, в то время как другие селективны по отношению только к определенным анионами или катионам. Селективность определяется особой областью ионного канала, называемой селективный фильтр.
Фильтр обеспечивает частичную дегидратацию ионов, проходящих через канал, что делает их транспорт энергетически более выгодным по сравнению с другими ионами одинаковой величины. Процесс открытия и закрытия ионных каналов называется воротным механизмом (гейтинг) и регулируется разнообразными факторами. Например, ионный канал может открываться и закрываться под действием электрического заряда, механических усилий или температуры. Направление потока ионов через канал зависит от электрохимического градиента того или иного иона, а его транспорт связан с прохождением электрического тока через мембрану. Выяснено строение некоторых канальных белков, и предложены модели, объясняющие их селективность, быструю скорость транспорта и контроль воротного механизма.
Переносчики и насосы представляют собой селективные белки, которые отличаются от каналов механизмами переноса. Транспортные белки способны попеременно принимать две основные конформации: в одной конформации они способны связывать переносимый метаболит на одной стороне мембраны, а находясь в другой, высвобождать его с противоположной стороны. Для осуществления процесса транспорта переносчики и насосы используют различные источники энергии. Переносчики расходуют энергию электрохимического градиента по обеим сторонам мембраны. Напротив, насосы используют энергию АТФ или внешние источники (например, свет).
Трансмембранный электрохимический градиент устанавливается за счет взаимодействия различных транспортных белков. Большая часть энергии клетки расходуется на установление ионного градиента по сторонам или плазматической мембраны или внутренних мембран клетки. Например, за счет насоса Na+/К+-АТФазы создается трансмембранный градиент ионов Na+ и К+. Энергия электрохимических трансмембранных градиентов используется каналами и переносчиками для выполнения работы.
Эти градиенты, например, используются потенциал-зависимыми ионными каналами для генерации электрических зарядов, или другими типами ионных каналов для активации внутриклеточных систем передачи сигнала, а также для контроля клеточного объема или управления транспортом жидкостей и электролитов. Вместе с тем, переносчики используют энергию электрохимического градиента одного метаболита для транспорта другого метаболита против градиента его концентрации.
Мутации в генах, кодирующих белки ионного канала, переносчика или насоса, вызывают различные функциональные дефекты. Нарушения функции ионных каналов часто проявляются в нейронах и мышечных клетках и служат причиной развития таких заболеваний, как эпилепсия, атаксия, миотонии и сердечные аритмии. К числу болезней при нарушении ионных каналов, затрагивающих другие органы, относятся кистозный фиброз, синдром Бартера, расстройства секреции инсулина, образование почечных камней и остеопетроз. Исследования характера нарушений ионных каналов при мутациях позволяют понять особенности их функционирования и физиологическую роль.
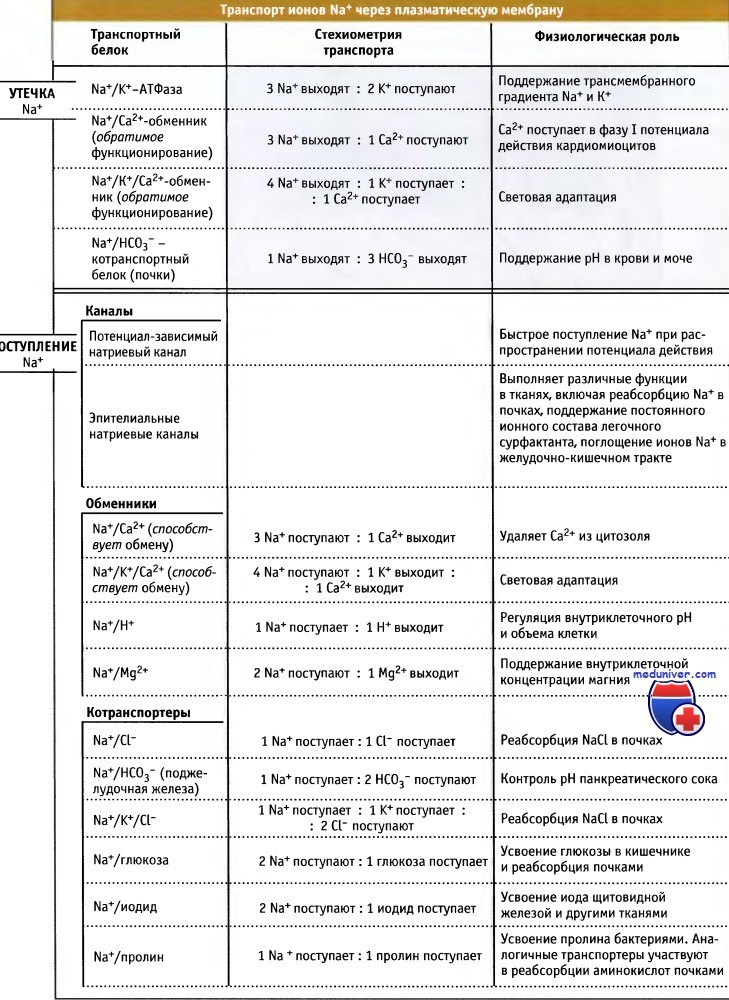
Na+/K+-АТФаза поддерживает градиент Na+ по сторонам плазматической мембраны, удаляя из клетки ионы Na+.
Транспорт ионов в клетку в направлении градиента обеспечивается Na+-каналами.
Некоторые переносчики используют высвобождающуюся при этом энергию для транспорта других метаболитов против градиента концентрации.