Перспективы изучения обмена и адресования белков в клетке
Большая часть статей на сайте посвящена объяснению того, каким образом белки переносятся через мембрану эндоплазматического ретикулума (ЭПР). В основном это связано с тем, что транспортная функция ЭПР изучена лучше всего. Достаточно подробно исследован основной путь транслокации секреторных белков. Однако вопросы, связанные с транслокацией более сложных субстратов, остаются неисследованными, в особенности это касается интеграции мембранных белков.
Каким образом ориентируются трансмембранные домены и что определяет их интеграцию в мембрану? Какую роль играет взаимодействие между ТМ доменами, которое происходит до момента их интеграции? Наряду с этим, становится ясно, что транслокация представляет собой регулируемый процесс, при котором эффективность позиционирования, переноса и интеграции белка, а также необходимые для этого факторы, существенно зависят от субстрата и от условий, в которых находится клетка. Пока мы не понимаем, как регулируется транслокация и каким образом клетка осуществляет этот процесс в соответствии со своими запросами.
Хотя в настоящее время выяснена структура канала транслокации, способ мобилизации и воротный механизм транслокона еще исследуются. Каким образом узнавание сигнальной последовательности приводит к структурным изменениям канала? Обладает ли структура канала гибкостью, достаточной для переориентации переносимой цепи, или для обеспечения накопления нескольких трансмембранных доменов? Что происходит со структурой канала по окончании транслокации? В какое время и каким образом белки, находящиеся поблизости от транслокона, связываются с ним?
Еще менее изучен вопрос, как ЭПР осуществляет сворачивание белков, а также узнает и удаляет белки с неправильной структурой. Почти совершенно не исследован наиболее фундаментальный аспект процесса, каким образом органел-ла «чувствует» правильно и неправильно собранный белок Многое предстоит выяснить о том, как шапероны взаимодействуют с белками. Необходимо определить, насколько тесно взаимодействуют между собой многочисленные системы шаперонов ЭПР и обладают ли они разными или перекрывающимися функциями. Также остается неясным, каким образом клетка принимает решение о деградации белка, обладающего неправильной структурой.
Существование механизма, ограничивающего длительность его взаимодействия с шаперонами, представляется привлекательным, однако какие-либо предположения о том, каким образом он может работать, отсутствуют. Наконец, неизвестно, как белок, отобранный для деградации, направляется обратно в канал и в цитозоль. Это же относится к механизму, посредством которого канал может открываться изнутри.
Еще менее изучена роль ЭПР в синтезе и сортировке липидов. Отчасти это связано с тем, что, по сравнению с белками, липиды труднее поддаются анализу, поскольку с ними гораздо сложнее манипулировать экспериментально. Дискуссионным остается основной вопрос, связанный с мембранными липидами: каким образом осуществляется их адресный транспорт из ЭПР в соответствующие мембраны. Обнаружение физического взаимодействия ЭПР с другими органеллами, например с мембранами митохондрий, предполагает существование возможного механизма переноса. Однако конкретная роль таких контактов не установлена. Также остается неясным, каким образом ЭПР регулирует синтез отдельных фосфолипидов.
Наконец, неизвестны механизмы, управляющие динамикой ЭПР в целом. Каким образом органелла сохраняет свою характерную форму? Как поддерживаются субкомпартменты? Как осуществляется связь ЭПР с цитоскелетом и как он перемещается в клетке? Чем определяются размеры ЭПР и каким образом он расширяется? Существование отклика неструктурированных белков свидетельствует о взаимосвязи между ЭПР и ядром. Возможно, что наличие такой взаимосвязи поможет ответить на некоторые из перечисленных особенностей ЭПР. Наличие контактов между ЭПР и другими компартментами клетки позволяет предполагать, что между ними возможен обмен, однако, каким конкретным образом это может происходить, остается неизвестным.
В клетке содержится много разных органелл, окруженных мембранами, некоторые из которых импортируют белки прямо из цитозоля. Митохондрии, хлоропласта и пероксисомы импортируют необходимые белки. Эндоплазматический ретикулум (ЭПР) также импортирует белки из цитозоля, однако большинство этих белков транспортируется дальше, или секретируется, или функционирует в органелл ах и мембранах, которые не могут импортировать белки сами. К их числу относятся плазматическая мембрана и органеллы секреторного или эндоцитозного пути.
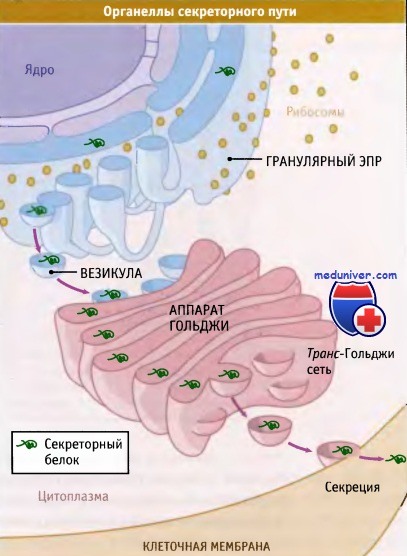
Секреция белков начинается с их адресования и транслокации через мембрану гранулярного эндоплазматического ретикулума.
После формирования нативной структуры и посттрансляционных модификаций белки выходят из ЭПР в везикулах, связывающихся с аппаратом Гольджи.
Большая часть белков переносится из аппарата Гольджи к клеточной поверхности в секреторных везикулах.
Белки, импортируемые в органеллы, идентифицируются с помощью сигнальной последовательности, представляющей собой короткую цепочку аминокислот, обычно расположенную на N-конце цепи. Последовательности, соответствующие разным органеллам, различаются по длине и по составу. Последовательности ЭПР содержат около двадцати аминокислот и характеризуются протяженным участком, состоящим из гидрофобных остатков. Близкими размерами обладают сигнальные последовательности митохондрий. Однако они содержат чередующиеся группы гидрофобных и заряженных аминокислот. Поэтому, когда сигнальная последовательность принимает форму а спирали, одна ее сторона становится гидрофобной, а другая заряженной.
Сигнальная последовательность пероксисом обычно состоит только из трех аминокислотных остатков. Во всех случаях белки идентифицируются и позиционируются больше не в соответствии с составом сигнальных последовательностей, а на основании их физических свойств.
Каждый тип сигнальной последовательности связывается со специфическим рецептором, который направляет белок к определенной органелле. Когда белок достигает органеллы, он транслоцируется внутрь нее по мембранному каналу. От момента распознавания сигнальной последовательности, при синтезе белка или после его завершения, зависит тип транслокации. Котрансляционная транслокация происходит в случаях, когда сигнальная последовательность узнается в процессе синтеза белка. При этом, рибосома, на которой синтезируется белок, связывается с мембраной, и образующийся белок переносится в канал транслокации. Посттрансляционная транслокация происходит, когда сигнальная последовательность узнается после завершения синтеза белка.
Большинство белков попадают в ЭПР путем котранс-ляционной транслокации. Сразу после выхода сигнальной последовательности с рибосомы с ней связывается частица, распознающая сигнал (SRP). Рибосома и растущий белок позиционируются на мембране ЭПР за счет взаимодействия между SRP и ее рецептором. Затем рибосома вместе с растущей цепью занимают канал, по которому белок проходит через мембрану. Сердцевину канала образует комплекс Sec61, вокруг — большая группа белков, участвующих в транслокации. Все вместе это называется транслоконом. При взаимодействии с сигнальной последовательностью канал открывается, что позволяет цепи войти в него таким образом, который исключает транспорт других молекул через мембрану ЭПР. Трансляция продолжается до тех пор, пока рибосома остается связанной с каналом, и белок транспортируется по нему в люмен.
Интеграция белка в мембрану ЭПР начинается в тот момент, когда трансмембранный домен начинает транслироваться и входить в канал. Поскольку трансмембранные домены обладают гидрофобностью, они узнаются каналом, и проходят через его стенки в липидный бислой мембраны. После узнавания трансмембранного домена транспорт образующегося белка через мембрану прекращается. Трансляция, однако, продолжается, что приводит в выходу дальнейший участков полипептидной цепи в цитозоль. Для полной интеграции мембранных белков с несколькими трансмембранными доменами, вероятно, необходимо многократное открытие и закрытие канала.
Процесс интеграции мембранных белков осложняется необходимостью их определенной ориентации в мембране. Очевид но, что она зависит от свойств трансмембранных доменов белков. Однако не ясно, как домены должны взаимодействовать с каналом, мембранными липидами, или друг с другом, чтобы определить ориентацию белков.
Некоторые белки транслируются в ЭПР посттрансляционно. Связывание шаперонов в цитоплазме предотвращает сворачивание этих белков, и пока они находятся в процессе адресования, поддерживает их в состоянии, готовом для транслокации. Адресование обеспечивается при связывании их сигнальных последовательностей с Sec6l в канале транслокации. Это тот же канал, который используется при котрансляционной транслокации, однако содержащий четыре дополнительных белка, включая Sec62 и Sec63. Вновь образованные белки проходят через канал за счет энергии, высвобождающейся при гидролизе АТФ белком BiP в люмене ЭПР. BiP использует гидролиз АТФ для связывания и высвобождения белка на конце канала со стороны люмена. Молекулы BiP остаются в связанном состоянии достаточно долго и действуют как храповик, продвигающий белок в люмен.
При транслокации в ЭПР многие белки подвергаются ковалентной модификации. Обычно сигнальная последовательность удаляется пептидазой вскоре после того, как белок вошел в канал. Остальные участки молекулы транслоцируемого белка часто модифицируются после того, как они прошли в люмен. К белку могут добавляться остатки сахаров олигосахаридтрансферазой, или за счет действия протеиндисульфидизомеразы (ПДИ) в молекуле могут образоваться дисульфидные связи. Некоторые полностью транслоцированные белки расщепляются рядом с С-концевым участком и присоединяются к гликозилфос-фатидилинозитолу (ГФИ), фосфолипиду, который связывает их с мембраной.
После попадания в люмен белки начинают приобретать нативную структуру. Это обеспечивается большим количеством разнообразных шаперонов. BiP и Grp94 непосредственно взаимодействуют с неструктурированными белками. Кальнексин и кальретикулин связываются с сахарными остатками, которые добавляются к белкам при транслокации. Они также участвуют в процессе добавления глюкозного остатка к полипептидной цепи. Наличие этого остатка позволяет судить о том, насколько правильной нативной структурой обладает белок ПДИ участвует в перегруппировке дисульфидных связей, которая сопровождает структурирование белка. После приобретения белками правильной нативной структуры они уже не реагируют с шаперонами и получают возможность выйти из ЭПР в аппарат Гольджи Если несколько попыток образования белком правильной нативной структуры оказались безуспешными или белок не может образовать комплекс с другими белками, то он возвращается в канал и назад в цитозоль за счет ретроградной транслокации. Когда он попадает в цитозоль, то подвергается деградации в протеосомах.
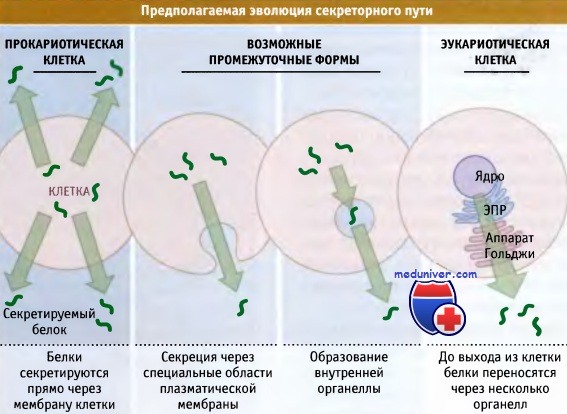
Органеллы секреторного пути могли возникнуть при интернализации участка плазматической мембраны,
обладающей функцией секреции белка.
В клетках эукариот белки непосредственно не секретируются через плазматическую мембрану,
а транспортируются в люмен ЭПР, который по составу напоминает внешнее окружение клетки.
Накопление в ЭПР больших количеств белков, не имеющих нативной структуры, вызывает отклик неструктурированных белков. Это цепь сигналов от ЭПР к ядру, которые обеспечивают дополнительный синтез шаперонов. Сигнал опосредуется трансмембранными белками, резидентными компонентами ЭПР, которые узнают присутствие неструктурированных белков, не связанных с BiP. При этом запускаются события, в результате которых изменяется экспрессия генов, и ЭПР получает возможность преодолеть последствия избытка неструктурированных белков. В-случае высших эукариот, при продолжающемся состоянии стресса, эти же процессы вызывают клеточную гибель.
Импорт белков в митохондрии и хлоропласты происходит после окончания трансляции. Для обеих органелл характерно существование двух мембран. Белки могут быть локализованы в одной из мембран, в межмембранном пространстве или внутри органеллы. Каждая мембрана имеет свой транслокон. У митохондрий они носят название ТОМ для наружной мембраны и TIM для внутренней мембраны, а у хлоропластов ТОС и TIC соответственно. Сигнальные последовательности узнаются транслоконом наружной мембраны. Транслоконы внутренней и наружной мембран связаны между собой, так что импортируемые белки переносятся между ними непосредственно. Белки митохондрий могут переноситься через обе мембраны и затем при узнавании отдельной сигнальной последовательности снова направляться к внутренней мембране.
В транслокации белков хлоропластов через мембрану внутренней органеллы, называемой тилакоидом, также участвует особая сигнальная последовательность.
Транслокация белов в митохондрии происходит за счет электрохимического градиента через внутреннюю мембрану и сил взаимодействия между импортируемыми белками и шаперонами в матриксе митохондрий. За счет чего происходит транслокация в хлоропласты, остается неясным. Мы также не знаем, каким образом происходит интеграция белков в мембраны этих органелл.
Транспорт белков в пероксисомы происходит посттрансляционно, но отличается от их транслокации в другие органеллы. При импорте в пероксисомы белок проходит через одну мембрану, и процесс начинается после того, как он приобрел в цитозоле нативную структуру. Сигнальная последовательность для пероксисом узнается в цитозоле при участии белков, которые остаются связанными с субстратом в момент его транслокации. Белки-переносчики диссоциируют только после того, как они оказались внутри органеллы, и возвращаются в цитозоль для дальнейшего использования. Происхождение белков мембраны пероксисом остается неизвестным.
Наряду с импортом, созреванием и распределением белков, ЭПР выполняет в клетке еще несколько функций. Его функции отражаются в структуре. Транслокация и созревание белков происходят в гранулярном ЭПР, покрытом рибосомами. Гладкий ЭПР отличается от гранулярного. Обычно он представляет собой трубочки, организованные в динамичную сеть, разветвленную по всему цитозолю. Гладкий ЭПР часто связан с элементами цитоскелета и контактирует с другими мембранами клетки. К числу его функций относится синтез липидов для всех клеточных мембран. Липиды должны каким-то образом транспортироваться из ЭПР в другие мембраны, однако неизвестно, как это происходит. Возможно, что это происходит в точках контакта гладкого ЭПР с другими мембранами. ЭПР также служит резервуаром внутриклеточного кальция. Кальций выходит в ответ на получаемый клеткой сигнал, и после выполнения своих функций закачивается назад в органеллу.
В специализированных клетках в гладком ЭПР могут также синтезироваться жирорастворимые гормоны или обезвреживаться потенциально опасные для клетки химические соединения. В клетках, специализированных для выполнения какой-либо функции, осуществляемой за счет ЭПР, например секретирующих много белка, или образующих стероидные гормоны, может дополнительно образовываться гранулярный или гладкий ЭПР, и занимать в них большую часть цитозоля. В таких высокоспециализированных клетках как скелетные мышцы, ЭПР также чрезвычайно высокоспециализирован по своему составу и строению. Саркоплазматический ретикулум, который представляет собой специализированный гладкий ЭПР, обернут вокруг саркомеров скелетных мышц и приспособлен для доставки ионов кальция, стимулирующих мышечное сокращение.