• Митохондрии обладают внутренней и внешней мембранами, в каждой из которых присутствует транслокационный комплекс
• Импорт белков в митохондрии происходит после завершения трансляции
• Митохондриальная сигнальная последовательность узнается рецептором, расположенным на наружной мембране
• Комплексы ТОМ и TIM физически связаны между собой и при импорте белки непосредственно переходят от одного к другому
• Импорт белков обеспечивается энергией за счет Hsp70 матрикса митохондрий и мембранного потенциала внутренней мембраны
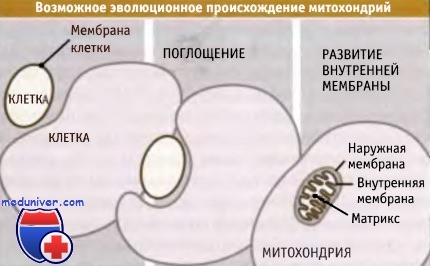
Считается, что митохондрии произошли из клеток прокариот, захваченных другими, более крупными клетками, что привело к развитию между ними симбиотических отношений. В результате таких эволюционных событий, в отличие от других органелл, митохондрия оказалось окруженной двумя мембранами, а не одной. Белки, предназначенные для внутренней части митохондрии (матрикса митохондрий), должны пройти через обе мембраны и через промежуток между ними (межмембранное пространство). Каждая мембрана пронизана специфическим мультиферментным комплексом, который включает канал транслокации.
Комплекс ТОМ (транслоказа внешней мембраны) пронизывает внешнюю мембрану, a TIM (транслоказа внутренней мембраны) — внутреннюю. В специфических точках между мембранами оба комплекса находятся в контакте, но они способны действовать независимо.
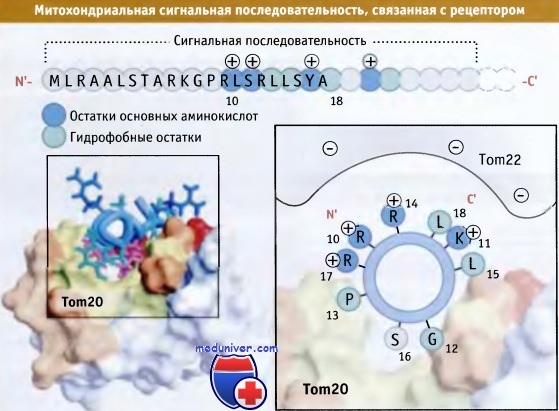
Митохондриальные белки переносятся к мембране после окончания трансляции. При этом узнается N-концевая сигнальная последовательность, длина которой обычно составляет 20-50 аминокислот. Эта последовательность обогащена как основными (положительно заряженными), так и гидрофобными аминокислотами. Эти остатки образуют амфипатические спирали, заряженные группы которых размещены с одной стороны, а гидрофобные остатки — с другой.
Сигнальная последовательность митохондриального белка вначале узнается Tom20, интегральным мембранным белком, который является компонентом ТОМ-комплекса. Tom20 связывается с гидрофобной стороной сигнальной последовательности, находящейся в мелкой бороздке (в отличие от глубокой и гибкой бороздки, которая используется SRP при таргетинге в ЭПР).
Как показывает рисунок ниже, взаимодействие между сигнальной последовательностью и Tom20 носит слабый характер и позиционирует положительно заряженные остатки в сигнальной последовательности вне связывающего кармана. Затем взаимодействие сигнальной последовательности с ТОМ-комплексом укрепляется вторым белком, Tom22. Tom22 содержит кислый цитозольный домен, который взаимодействует с остатками основных аминокислот в составе сигнальной последовательности. В основном в результате этих двух взаимодействий белок входит в контакт с комплексом ТОМ. При таргетинге митохондриальные белки находятся в несвернутом состоянии и подготавливаются к транслокации за счет связи с hsp70, находящимся в цитозоле.
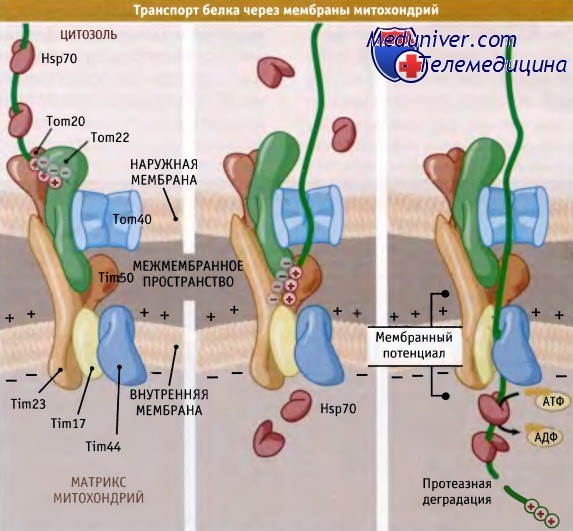
Рисунок ниже иллюстрирует транспорт митохондриального белка с участием комплексов ТОМ и TIM. Наряду с белками Tom20 и Tom22, комплекс ТОМ содержит и другие белки. Из них в образовании канала, вероятно, принимает участие белок Тот40. До настоящего времени остаются не вполне понятными ни механизм узнавания транслоцируемого белка в канале, ни воротный механизм канала. Однако после прохода в канал основная сигнальная последовательность субстрата за счет сил электростатического притяжения входит в контакт в межмембранном пространстве с кислыми доменами белков Tom22 и Tim23.
Последний входит в комплекс TIM. Близость Tom22 и Tim23 обеспечивает прохождение белка между этими комплексами, минуя выход в межмембранное пространство. Существуют данные, свидетельствующие о том, что N-концевой участок Tim23 может быть ингегрирован во внешнюю мембрану. Это приводит к тесной ассоциации комплексов TIM и ТОМ и обеспечивает эффективный перенос белковой цепи от одного комплекса к другому. Предполагается также, что важная роль в транспорте предшественников белков от комплекса ТОМ в канал TIM23 принадлежит белку Tim50.
После причаливания цепи в комплексе TIM (который образуется в основном белками Tim23, Tim17 и Tim44; из них два первых, вероятно, образуют канал) она продвигается в матрикс под действием двух сил. Белок mtHsp70, являющийся гомологом в матриксе белка, Hsp70, связывается с каналом посредством взаимодействия с Tim44. Так же как и при посттрансляционной транслокации в ЭПР, mtH-sp70 связывает субстрат по мере его выхода из канала и действует как храповой механизм, или как мотор, а возможно, осуществляет обе функции. Многое здесь зависит от транслоцируемого субстрата.
Транслокации также способствует электрохимический потенциал внутренней мембраны, вероятно, за счет взаимодействия с положительно заряженной сигнальной последовательностью. Потенциал направлен поперек мембраны, что благоприятствует движению положительных зарядов в направлении матрикса. Пока, однако, неизвестно, каким образом потенциал, действительно, способствует транслокации. После входа в матрикс большинство сигнальных последовательностей отщепляются растворимой митохондриальной процессирующей протеазой (МРР).
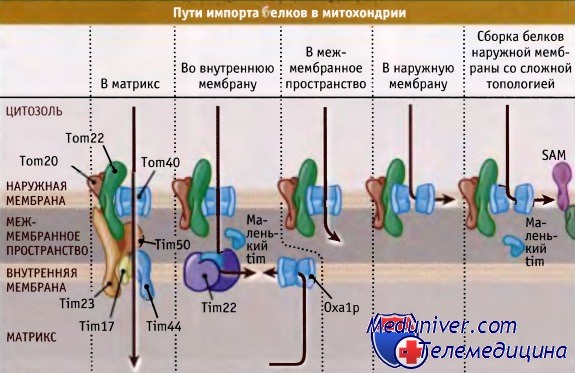
Так же как и в случае белков, интегрированных в мембрану ЭПР, белки митохондриальной мембраны часто позиционируются с участием внутренней неотщеп-ляемой сигнальной последовательности. Эта последовательность также содержит инструкции по интеграции в виде отдельных участков гидрофобных аминокислот. Каким образом узнаются и интегрируются в одну из двух мембран трансмембранные домены, в общем, неизвестно. Белки, которым необходимо интегрироваться во внутреннюю мембрану, позиционируются в альтернативном транслоконе, состоящем из Tim22, Tim54 и Tim18. Транспорту белка из комплекса ТОМ в этот альтернативный транслокон способствуют небольшие белки, которые называются «Маленькие Тимы», и которые находятся в межмембранном пространстве. Последовательность событий представлена на рисунке ниже. Подробнее об этих процессах почти ничего не известно.
Белки внутренней мембраны, независимо от их кодирования в ядре или в митохондрии, интегрируются с участием белка внутренней мембраны Оха1р. Белки внутренней мембраны, кодируемые в ядре, вначале транслоцируются через обе мембраны в матрикс. Затем от белка отщепляется сигнальная последовательность, за счет которой происходит импорт, и еще одна последовательность направляет белок обратно к внутренней мембране, где она интегрируется с участием Оха1р. Белки, кодируемые митохондриальным геномом, синтезируются рибосомами матрикса, и затем непосредственно интегрируются во внутреннюю мембрану с участием Оха1р. Детали этого процесса пока точно не выяснены.
Идентифицирован еще один транслокон митохондрий, названный «комплекс SAM» (sorting and assembly machinery of the outer membrane complex; сортировочно-сборочный комплекс наружной мембраны). Хотя состав и механизм действия этого комплекса в деталях неизвестны, он, вероятно, благоприятствует транслокации и интеграции белков внешней мембраны, которые пронизывают мембрану b-структурами, а не более обычными а-спиралями. Так же как и комплекс Tim22, для транспорта субстратов из ТОМ канала, комплекс SAM использует «Маленьких Тимов».
Особый интерес представляет вопрос, как белки внутренней мембраны проходят через наружную мембрану и при этом не интегрируются в нее. Такая специфичность может объясняться различиями между трансмембранными доменами белков внутренней и наружной мембран, которые делают белки внутренней мембраны неузнаваемыми комплексом интеграции ТОМ. В то же время, не исключено, что комплекс TIM может узнавать белки внутренней мембраны до того, как они интегрируются во внешнюю мембрану комплексом ТОМ. Пока еще не совсем понятны воротные механизмы, регуляции и координации комплексов ТОМ и TIM. Хотя многие выполняемые ими функции аналогичны функциям транслокона ЭПР, остается неясным, насколько близки механизмы их функционирования.
При формировании спиральной структуры последовательность митохондриального сигнала образует поверхности с основными и гидрофобными свойствами.
Последовательность аминокислот, соответствующая митохондриальному сигналу,
представлена наверху, и та же последовательность, свернутая в спираль, показана справа.
Гидрофобный участок спирали взаимодействует с гидрофобной щелью Tom20; остатки положительно заряженных аминокислот взаимодействуют с кислым доменом Tom22.
Слева показана структура Tom20, связанная с сигнальной последовательностью; гидрофобная поверхность белка выделена желтым цветом.
Белок, предназначенный для транслокации в матрикс митохондрии,
узнается на поверхности наружной мембраны и затем проходит между каналами в наружной и внутренней мембранах.
Когда белок достигает матрикса, сигнальная последовательность удаляется.
Необходимая для транспорта энергия поставляется за счет гидролиза АТФ шаперонами матрикса и за счет мембранного потенциала на внутренней мембране.
Белок Tim23 интегрирован в обе мембраны и связывает между собой каналы.
Для импорта белков в различные отсеки митохондрий требуются различные пути.
Во многих из них может участвовать комплекс ТОМ, однако для комплексов внутренней мембраны характерна специализация.
Существуют два пути, посредством которых белки могут достигать внутренней мембраны.
Некоторые белки используют комбинированный путь: вначале они транспортируются в матрикс, а потом поступают назад в мембрану.