• Рецепторы для импорта белков в ядро представляют собой цитоплазматические белки, которые связываются с NLS транспортируемых белков
• Рецепторы ядерного импорта представляют собой часть большого семейства белков, которые называются кариоферины
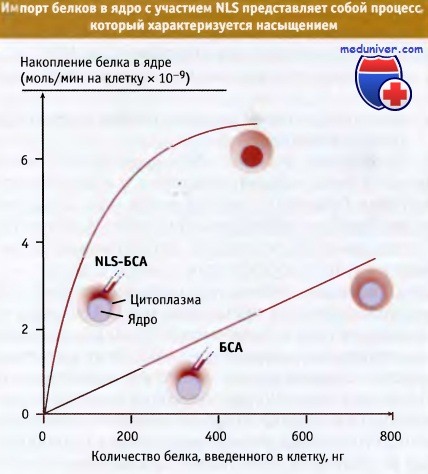
Импорт белков в ядро представляет собой рецепторный процесс. На основании экспериментов, показавших, что процесс импорта белков характеризуется насыщением, предположили участие в нем рецепторов, узнающих сигнал ядерной локализации.
В экспериментах измеряли кинетику ядерного импорта в ооциты Xenopus. Короткие полипептиды, содержащие NLS, с помощью химических кросслинкеров сшивали с неядерным белком, бычьим сывороточным альбумином (БСА). Когда комплекс NLS-БСА вводили в цитоплазму ооцитов, он проникал в ядро, в то время как индивидуальный БСА оставался в цитоплазме. По мере увеличения концентрации NLS-БСА, процесс накопления белка в ядре характеризуется кривой с насыщением.
Это согласуется с точкой зрения о наличии ограничивающего фактора (или факторов), узнающих NLS и осуществляющих импорт, что, в свою очередь, предполагает существование рецепторов, узнающих NLS-содержащие белки.
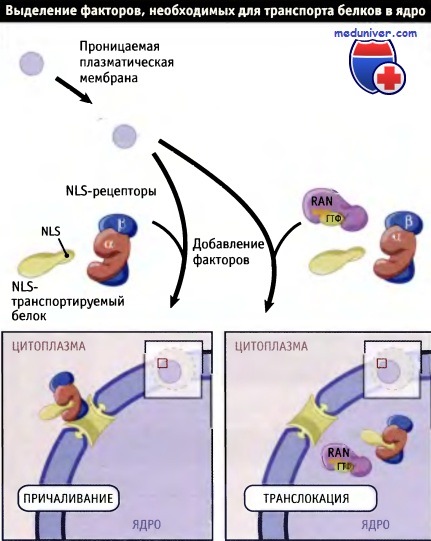
Сейчас мы знаем, что рецепторы NLS представляют собой растворимые цитоплазматические белки. Некоторые рецепторы были впервые идентифицированы с помощью метода, который in vitro воспроизводит события в клетке и позволяет выделить цитозольные факторы, обеспечивающие процесс импорта белков. Клетки обрабатывали детергентом дигитонином, чтобы сделать плазматическую мембрану проницаемой.
Такая обработка позволяет белкам цитоплазмы выходить через отверстия в мембране, однако ядерная оболочка при этом не меняется. Когда к проницаемым клеткам добавляли NLS-содержащие белки, они поступали в клетку через отверстия в плазматической мембране, но не проникали в ядро, если к ним не добавляли экстракт цитоплазмы.
После введения в цитоплазму ооцита комплекса NLS-БСА,
импорт его в ядро характеризуется насыщением.
Из экстракта цитоплазмы были выделены два белка — один белок связывался с транспортируемым белком и ядерной порой, а другой участвовал в транслокации. Таким образом, импорт ядерных белков происходит в два этапа:
• NLS-зависимое причаливание белка к ядерной поре с последующей
• транслокацией его в нуклеоплазму.
Белок, обеспечивающий причаливание, был идентифицирован как NLS-рецептор. Этот рецептор присоединяется к транспортируемому белку (карго) через NPS, переносит транспортируемый белок в ядро и затем возвращается в цитоплазму для следующего цикла транспорта. Транслокацию через ЯПК обеспечивает Ran-ГТФаза.
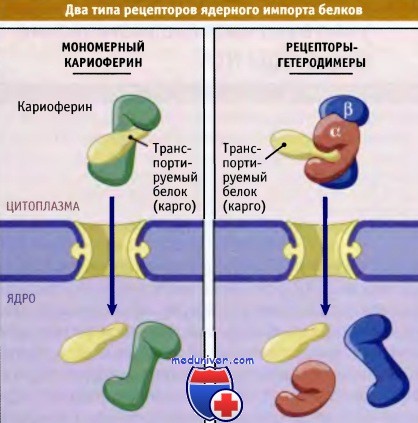
Идентифицированы два типа рецепторов NLS:
• рецепторы, непосредственно связывающиеся с порой, и
• рецепторы, которым для причаливания белка к поре требуется адаптер.
Простейший рецептор NPS представляет собой молекулу белка, которая связывается с NLS-транспортируемым белком и с ядерной порой. Эти рецепторы очень напоминают импортен b. Напротив, у гетеродимерных рецепторов одна субъединица связывается с транспортируемым белком, а другая — с нуклеопоринами. Первый идентифицированный NLS рецептор состоял из импортинов а и b. Импортен а представляет собой белок массой 55-60 кДа, который непосредственно связывается с NLS.
Этот импортен является специфическим адаптером классического NLS. Белки с таким NLS преобладают и используют импортен а как адаптер, позволяя большому количеству транспортируемых белков переноситься с импортином b. Последний представляет собой белок порядка 90 кДа, и вместе с импортином а удерживает белок в ядерной поре. Импортен а — только один из многих белков, которые узнают NLS и взаимодействуют с импортином b.
Для очистки цитоплазматических белков, необходимых для ядерного транспорта,
были использованы клетки с полупроницаемой мембраной.
В трехмерной структуре импортина а, связанного с NLS SV40, присутствуют два связывающих кармана для NLS. Это объясняет способность двойных и простых NLS взаимодействовать с одним и тем же рецептором. Третий лизиновый остаток в NLS участвует в создании трех критических контактов между аминокислотами и играет наиболее важную роль в связывании NLS. Это позволяет объяснить, почему даже столь незначительные вариации в первичной последовательности NLS, как замена лизина в третьем положении, полностью лишает белок функции.
После связывания с цитоплазматическими элементами ЯПК-комплексы кариоферин-транспортируемый белок проходят через канал поры, однако механизм этого процесса остается неизвестным. Для того чтобы выяснить, сколько последовательных контактов комплекса с нуклеопорином необходимо для транспорта и по какому механизму происходит этот процесс, необходимы дальнейшие исследования. Ниже рассматриваются две модели транспорта белков через пору.
Некоторые транспортируемые белки для импорта используют более одного рецептора ядерного транспорта. Наличие нескольких рецепторов может помочь обеспечить эффективный импорт и/или сформировать другой уровень регуляции. Более того, не все белки импортируются в ядро с участием кариоферинов. Некоторые белки, такие как сигнальные молекулы b-катенина, распределяются между любыми типами транспортеров и при импорте непосредственно взаимодействуют с нуклеопоринами.
Почему некоторые белки способны к транспорту без участия кариоферинов, неизвестно, однако это может быть вызвано необходимостью отдельной регулировки ядерного импорта, независимо от других транспортных процессов.
Некоторые белки вирусов приспособили собственные ядерные факторы для взаимодействия с нуклеопоринами и клеточными переносчиками. Например, вирусный белок Vpr, необходимый для инфицирования неделящихся клеток ВИЧ, связывает импортин а и нуклеопорины, что способствует импорту в ядро генетического материала вируса. Таким образом, Vpr имитирует участие импортина b в ядерном транспорте.
Хотя вначале считалось, что импортин b участвует только в процессе ядерного транспорта, в настоящее время известно, что он связан с регуляцией различных клеточных функций. Этот белок может взаимодействовать с карго и микротрубочками и, таким образом, обеспечивать продвижение ядерных белков вдоль микротрубочек к периферии ядра. Вместе с другими ядерными транспортными факторами импортин р играет роль в сборке митотического веретена и ЯПК, а также в формировании ядра после завершения митоза.
Многие белки поступают в ядро, непосредственно связываясь с кариоферинами, белками,
родственными импортину b (левая часть рисунка).
Вход в ядро других белков происходит с участием гетеродимера ар импортинов (показано справа)