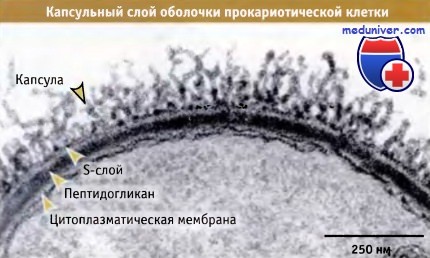
• У многих прокариот наружная поверхность содержит слой, обогащенный полисахаридами, который называется капсула или слизистый слой
• Предполагается, что капсула или слизистый слой предотвращает высыхание бактерий, обеспечивает их связывание с рецепторами клеток хозяина при попадании в организм, а также помогает бактериям избежать атаки со стороны его иммунной системы
• Формирование капсулы у Е. coli происходит по крайней мере по одному из четырех различных путей
• У многих прокариот, наряду с капсулой или вместо нее, существует S-слой. представляющий собой внешнее белковое покрытие обладающее кристаллическими свойствами
Большинство прокариот, если не все, поверх клеточной стенки образуют гликокаликс или капсулу. Термин гликокаликс приложим к клеткам как прокариот, так и эукариот; и используется для обозначения смеси внеклеточных полисахаридов и белков. В случае прокариотических клеток, между различными типами гликокаликса существуют довольно произвольные различия. Полисахариды, ковалентно связанные с клеткой, называются капсулой.
Непрочно связанные полисахариды, способные отщепляться от клетки, называются «слизистым слоем» или внеклеточными полисахаридами. Состав капсулы значительно варьирует, однако обычно в состав ее входят полисахариды, включая полиспирты и аминосахара, протеогликаны и гликопротеины. Толщина и степень пластичности этого слоя варьируют в зависимости от его химического состава. Полисахариды капсулы образуются при объединении моносахаридов в длинные цепи.
Поскольку любой моносахарид способен соединиться с другим моносахаридом, существует большое разнообразие капсулярных полисахаридов. Серологические свойства капсулярных ингредиентов можно использовать для выяснения отличий ее состава у родственных организмов. Например, у Е. coli описано более 80 различных капсулярных полисахаридов или К-антигенов. Известно, что штаммы, экспрессирующие некоторые К-антигены, связаны с определенными инфекциями.
У большинства прокариот капсула представляет собой наружный слой клеточной оболочки.
На этой микрофотографии представлена оболочка Bacillus anthracis.
Капсульный слой выполняет несколько функций, включая защиту клеток от высыхания, фагоцитов, детергентов и бактериофагов. Защита от высыхания играет важную роль в переносе инкапсулированных микроорганизмов от одного хозяина к другому. Капсула выполняет важную роль, обеспечивая адгезию клеток к тканям хозяина и к внешним поверхностям. Капсулы также обеспечивают связывание с другими прокариотами, что приводит к образованию биопленки. Например, Pseudomonas aeruginosa продуцирует огромные количества альгинатов, которые способствуют образованию биопленки в легких.
Такая прочная пленка из альгината также способствует защите патогенных микробов от антибиотиков и защитных систем организма хозяина.
Капсула играет важную роль не только при инфицировании организма хозяина патогенным микроорганизмом Р. aeruginosa. Например, основным вирулентным фактором Bacillus anthracis, вызывающего сибирскую язву, оказалась капсула. Капсула В. anthracis состоит из полимера D-глутаминовой кислоты. Это единственная капсула прокариот, состоящая из пептидов. Материал капсулы продуцируется in vivo, и штаммы В. anthracis, неспособные синтезировать капсулу, характеризуются ослабленной патогенностью. Считается, что, поскольку материал капсулы В. anthracis обладает слабой иммуногенностью, она защищает патогенный микроорганизм от атаки иммунной системы организма хозяина. Капсула также оказывает ингибирующее действие на связывание белков комплемента, тем самым позволяя микробу существовать в системе кровообращения хозяина.
Капсула также является основным фактором вирулентности Mycobacterium tuberculosis. У Mycobacterium капсула обеспечивает связывание с макрофагами, что служит критическим шагом в проникновении бактерии в организм хозяина. Поскольку макрофаги являются фагоцитами, которые захватывают и разрушают микробы, последние выработали различные механизмы, позволяющие им выживать находясь внутри макрофагов. Для Mycobacterium капсула играет критическую роль в одном из таких механизмов.
Во-первых, она обеспечивает связывание с CR3 рецептором макрофагов, что вызывает сигнальный каскад, приводящий к «безопасному» пути инфекции. Также она играет важную роль в нейтрализации активных кислород-радикалов, присутствующих в макрофагах и способных разрушить микроб. Это помогает объяснить, каким образом энкапсулированные Mycobacteria избегают атаки иммунной ситемы и персистируют в организме хозяина, в то время как бескапсульные штаммы обладают меньшей вирулентностью. В действительности, бескапсульные штаммы Mycobacterium используются для изготовления вакцины, обеспечивающей иммунитет к тубекулезу.
Биогенез капсулы изучен на нескольких видах бактерий, однако, вероятно, лучше всего на E. coli. По генетическим и биохимическим критериям капсулы этой бактерии подразделяются на четыре группы. Особенно подробно исследован биогенез групп 1 и 2. Транспорт капсул этих групп к поверхности клетки, вероятно, происходит в местах тесного контакта между цитоплазматической и наружной мембранами. В транслокации капсул, относящихся к первой группе, участвует высокомолекулярный комплекс, который образует Wza, представляющий собой липопротеид наружной мембраны. Предполагается, что Wza, представляющий собой белок, формирующий цилиндр (b-barrel), вместе с Wzc образует канал, через который происходит секреция предшественников капсулы. Эта система секреции функционально и генетически близка к Usher белку наружной мембраны, РарС.
При участии белков KpsE и KpsD, компоненты капсул группы 2 мигрируют через наружную мембрану в обоих направлениях. Белок KpsE связывается с плазматической мембраной через свой N-концевой домен, в то время как большая часть белка находится в периплазматическом пространстве. Хотя KpsE не пронизывает наружную мембрану, С-концевой участок его молекулы связан с ней. Функционирование такого секреторного аппарата зависит от взаимной близости цитоплазматической и наружной мембран. Хотя функция белка KpsD до конца не выяснена, известно, что он представляет собой периплазматический белок, который, возможно, необходим для мобилизации белка KpsE на наружной мембране. Биогенез капсул, относящихся к группам 3 и 4, изучен недостаточно.
У многих бактерий и археев на наружной поверхности находится белковая структура, которая называется поверхностный слой (S-слой). Ее можно рассматривать как альтернативу капсуле или как дополнение к ней. S-слои образуются из одного белка или фрагментов гликопротеинов, которые в процессе самосборки формируют кристаллическую решетку. Структура белка не является консервативной, однако часто он содержит много остатков кислых и гидрофобных аминокислот. Иногда, главным образом у археев и грамположительных бактерий, обнаруживаются гликозилированные белки. Особенно характерно наличие множественных сайтов гликозилирования.
Гликозильные цепи состоят из 20-50 идентичных субъединиц, содержащих набор сахарных остатков. На рисунке ниже представлен препарат S-слоя, приготовленный с использованием метода «замораживания-травления» (видна поверхность, обладающая гексагональной симметрией, и поперечный срез через S-слой). Обычно толщина S-слоя составляет от 5 до 25 нм. Наружная поверхность его довольно гладкая, а внутренняя выглядит сморщенной в тех местах, где слой взаимодействует с более глубоко лежащими слоями. Сборка S-слоя находится под контролем клеточной оболочки, которая взаимодействует с такими молекулами, расположенными на поверхности, как теихоевые кислоты (у грамположительных бактерий) или липополисахариды (у грамотрицательных бактерий). Каким образом сборка и позиционирование S-слоя связаны с капсулой (в случае когда существуют два слоя), пока не ясно.
Если у археев пептидогликан или эквивалентный компонент отсутствует, то S-слой представляет собой основной компонент клеточной оболочки. Хотя белки S-слоя составляют значительную часть тотального клеточного белка, их функции пока не вполне выяснены. Многие штаммы прокариот, культивируемые в лаборатории, утратили способность формировать S-слой, однако прекрасно растут. Возможно, существует некий смысл не формировать этот слой в условиях, когда он не требуется.
У некоторых прокариот снаружи вместо капсульного слоя находится S-слой.
Слева: электронная микрофотография, иллюстрирующая гексагональную симметрию наружной поверхности S-слоя.
Справа: электронная микрофотография поперечного среза клеточной оболочки, содержащей S-слой.