• Во всех исследованных геномах метазоа обнаружены гены белков промежуточных филаментов
• Семейство генов белков промежуточных филаментов эволюционировало за счет дупликации и транслокаций с последующими дупликациями
• У человека насчитывается 70 генов, кодирующих белки промежуточных филаментов
• У человека гены кератинов расположены кластерами, а гены белков промежуточных филаментов, не относящиеся к кератинам, рассредоточены по геному
По мере того как становится доступной информация о структуре генома различных организмов, предпринимаются попытки выяснить эволюционную историю генов, кодирующих синтез белков промежуточных филаментов, которые обнаружены в геноме у множества организмов, от млекопитающих до примитивных Метазоа. По-видимому, существование промежуточных филаментов особенно характерно для подвижных многоклеточных организмов. Промежуточные филаменты цитоплазмы, очевидно, возникли как средство упрочения клеток и тканей многоклеточного организма и с возникновением в ходе эволюции внешних структур, защищающих клетки (таких как жесткий экзоскелет), не утратились.
У Метазоа известны две линии генов белков промежуточных филаментов. Одна из них представляет собой ламиноподобный домен (L), содержащий длинный спирализованный домен 1В, а другая характеризуется коротким спирализованным участком (S), подобно гену промежуточных цитоплпазматических филаментов у человека. Две эти формы рано дивер-гировали в процессе эволюции, после разделения первичноротых и вторичноротых беспозвоночных, включающих позвоночных (организмов со скелетом) и других хордовых, обладающих нотохордом (предшественником настоящего скелета).
У первичноротых обнаружена только одна L-форма, а у всех хордовых присутствуют обе формы, причем L-форма характерна для современных ламинов, а S-форма — для современных промежуточных филаментов цитоплазмы. Это подтверждает точку зрения о существовании ламиноподобного предшественника семейства генов промежуточных филаментов.
Белки промежуточных филаментов беспозвоночных и позвоночных обладают многими общими чертами. Это говорит о том, что эти черты были заложены в них до дивергенции первично- и вторичноротых. Например, некоторые белки промежуточных филаментов червя Caenorhabditis elegans, обладают свойствами, близкими к кератинам, что указывает на появление облигатных гетерополимерных взаимодействий белков уже на ранних этапах эволюции. Напротив, в геноме плодовой мушки Drosophila melanogaster находится два гена ламинов, но не обнаружено генов белков цитоплазматических филаментов: в ходе эволюции организмов, обладающих экзоскелетом, промежуточные филаменты оказались утраченными. Вместо них у Drosophila существует система микротрубочек, которые выполняют некоторые необходимые функции, связанные с упрочением клеток.
Увеличение числа генов белков промежуточных филаментов произошло одновременно с возрастанием эволюционной сложности хордовых. Например, у одного из наиболее примитивных хордовых, оболочника dona intestinalis имеется 5 генов белков промежуточных филаментов, в то время как у Branchiostoma, организма близого и к позвоночным и к беспозвоночным, их 13. У сростночелюстной рыбы Fugu ribripes этих генов более 40, а у Homo sapiens — 70.
Относительные положения консервативных интронов (показаны красными вертикальными линиями)
в генах типов I-V промежуточных филаментов человека (за исключением ряда вариабельных хвостовых интронов).
Расположение интронов в генах проливает свет на их эволюционное происхождение. Как показано на рисунке ниже, у большинства генов промежуточных филаментов имеется 3-7 интронов, которые занимают примерно одинаковое положение, и находятся в области, кодирующей стержневой домен. По меньшей мере 5 позиций для этих интронов в геноме человека и низших позвоночных (Fugu ribripes) совпадают. Различное расположение интронов в генах белков группы IV позволяет предполагать разное эволюционное происхождение нейрофиламентов и остальных промежуточных филаментов. Возможно, это связано с действием обратной транскриптазы, поскольку, например, некоторые интроны отсутствуют.
Считается, что псевдогены, у которых отсутствуют интроны, образовались в результате действия ретровирусов на ранних этапах развития (ретровирусы инфицируют клетки посредством своей геномной РНК, на которой с помощью обратной транскриптазы образуется ДНК, которая интегрируется в геном клетки-хозяина). Для некоторых генов промежуточных филаментов характерно присутствие одного или двух псевдогенов, однако для простых первичных кератинов К8 и К18 обнаружено очень большое количество псевдогенов: 33 для К8 и 62 для К18, и многих фрагментов, рассеянных по геному. Поскольку гены KRT8 и KRT18 транскрибируются на ранних этапах эмбриогегнеза, когда количество клеток невелико, любые транспозиции генетического материала происходящие с участием ретровируса, могут включаться в половые клетки, сохраняться в них, и, таким образом, фиксироваться в геноме.
Близкое расположение в геноме родственных генов позволяет высказать предположение о том, как они возникли. У человека два семейства кератиновых генов сгруппированы в двух локусах на хромосомах 12 и 17. Это позволяет предполагать, что они возникли в результате дупликации. Кластер генов типа I на хромосоме 17q21.2 содержит 27 из 28 функциональных генов типа I и пять псевдогенов. Гены занимают 970 кбаз, и могут транскрибироваться в том или ином направлении. Этот кластер кератиновых генов прерывается другим кластером из 32 генов, размером 330 кбаз которые кодируют КАР, или белки, ассоциированные с кератином. Хромосома 12q 13-13 содержит кластер типа II, состоящий из 27 функциональных генов (26 генов типа II и один ген типа I) и восьми псевдогенов размером 780 кбаз ДНК. В этом случае все гены транскрибируются в одном направлении. Единственный ген типа I локализован на 12q и представляет собой ген, кодирующий простой эмбриональный кератин эпителия К18, который расположен рядом с геном, кодирующим кератин К8.
Возможно ли реконструировать последовательность эволюционных событий для этой большой и гетерогенной семьи генов с учетом функциональные особенностей последних? Вполне возможно, что «приоритет» получили функциональные ламины, которые возникли на самых ранних этапах эволюции путем естественного отбора. Они служили прочным футляром для предохранения целостности длинных нитей ДНК. Для того чтобы развиться в гены белков промежуточных филаментов цитоплазмы, гены ла-минов должны были утратить последовательности ядерной локализации и заякоривания. У млекопитающих последовательности, кодирующие промежуточные филаменты, достаточно консервативны и, очевидно, дивергировали до образования видов.
В эволюции хордовых, по мере формирования сложности строения эпителия, разнообразие кератинов обеспечивалось дупликацией и транслокацией. Наиболее древними кератинами, которые дивергировали первыми, были кератины К8 (тип II) и К18 (тип I). Эмбриональный кератин К8 в большей степени связан с белками типа III, и дупликация, наряду с утратой доменов Н1/Н2, привела к кератину, неспособному к самостоятельной сборке, т. е. к кератину К18, или белку-предшественнику типа I. Затем, вероятно, при дупликации и транслокации, К18 отделился от пролиферативных кератинов типа I, и за счет более интенсивной дупликации у человека на хромосомах 12 и 17 амплифицировались оба локуса.
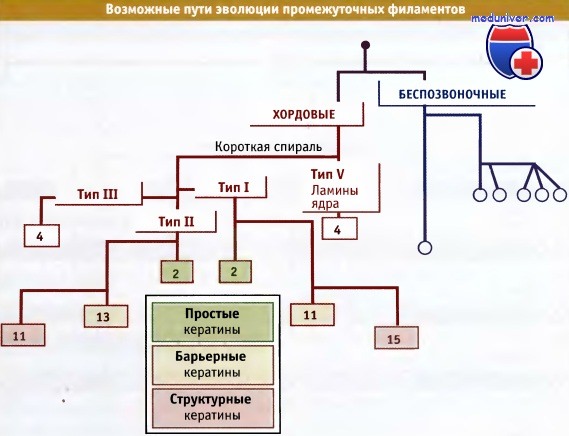
Трихоциты, или кератины волос, вероятно, являются последним продуктом эволюции кератинов, которая приводит к образованию меха, волос и когтей у млекопитающих, и «недавние» изменения между генами человека и больших человекообразных приматов показывают, что этот процесс еще не закончился. Изложенная схема эволюции белков промежуточных филаментов представлена на рисунке ниже. Поскольку гены белков типа IV обладают специфической структурой, их трудно разместить в рамках этой схемы, и поэтому они не показаны.
Наконец, в геноме всех известных растений и грибов не удалось обнаружить последовательностей, обладающих признаками свойственными генам, кодирующим белки промежуточных филаментов. Поэтому, хотя ламины являются эволюционно древними и необходимыми белками клеток млекопитающих, структурная организация ядра может формироваться и без их участия, при условии существования прочной клеточной стенки. У бактерий идентифицирован белок, по свойствам близкий к белку промежуточных филаментов. Это так называемый кресцентин, присутствующий в клетках Caulobacter crescentus. Аналогичный белок обнаружен у другой бактерии, Helicobacter pylori, являющейся представителем патогенной флоры желудка, присутствие которой связывают с развитием язвенной болезни.
В обоих случаях эти белки, вероятно, обеспечивают поддержание формы клетки и функционально гомологичны белкам промежуточных филаментов млекопитающих, хотя по последовательности аминокислот отличаются от них. Конвергентный характер эволюции подтверждает точку зрения о том, что у метазоа семейство генов белков промежуточных филаментов обширно и разнообразно потому, что эти белки играют важную роль в поддержании формы клеток в тканях.
Предполагаемое эволюционное происхождение генов белков промежуточных филаментов человека, относящихся к группам I—III и V.
Линии голубого цвета представлены только для ориентировки; цифрами указано количество генов в каждой группе.