• G-белки передают сигналы, регулируя активность множества внутриклеточных сигнальных белков, которые являются эффекторами
• Эффекторы обладают структурным и функциональным разнообразием
• У эффекторных белков не обнаружен общий домен связывания G-белка
• Эффекторные белки интегрируют сигналы от множества сигнальных путей, в которых участвуют G-белки
К эффекторам, активность которых регулируется G-белком, относятся ферменты, образующие или разрушающие вторичные мессенджеры (аденилатциклаза, фосфодиэстераза циклоГМФ, фосфолипаза С-β, фосфатидилинозитол-3-киназа), протеинкиназы, ионные каналы (К+, Са2+) и, вероятно, транспортные мембранные белки.
Эффекторы могут представлять собой белки, интегрированные в мембрану, или же растворимые белки, связывающие G-белки на поверхности мембраны. У эффекторных белков не обнаружено консервативного домена связывания G-белка или соответствующей последовательности, и большинство их представляют собой белки, обладающие близкими функциями, но которые не регулируются G-белками.
Таким образом, чувствительность к регуляции со стороны G-белков во многих семействах регуляторных белков возникла независимо.
Поскольку эффекторные белки могут реагировать на различные Gα- и Сβγ-субъединицы, они интегрируют сигналы от многочисленных сигнальных процессов, происходящих с участием G-белков. Различные субъединицы Gα или Gβγ могут проявлять противоположный или синергический эффект по отношению к данному эффектору.
Например, активность некоторых мембраносвязанных аденилатциклаз клеток млекопитающих стимулируется Gαs и ингибируется Gαi. Многие эффекторы регулируются другими аллостерическими лигандами (например, липидами, калмодулином) и фосфорилированием, что обеспечивает еще большую степень интеграции информации.
Обычно существуют множественные изоформы эффекторов, причем каждая изоформа регулируется по-разному, что еще добавляет сложности к организации регуляторной сети с участием G-белка. Например, активность некоторых изоформ аденилатциклазы под действием Gβγ стимулируется, в то время как активность других ингибируется. Под действием представителей семейства Gαq и Gβγ все фосфолипазы С-β стимулируются, однако степень выраженности и максимум проявления эффекта этих сигналов для четырех изоформ PLC-β сильно отличаются.
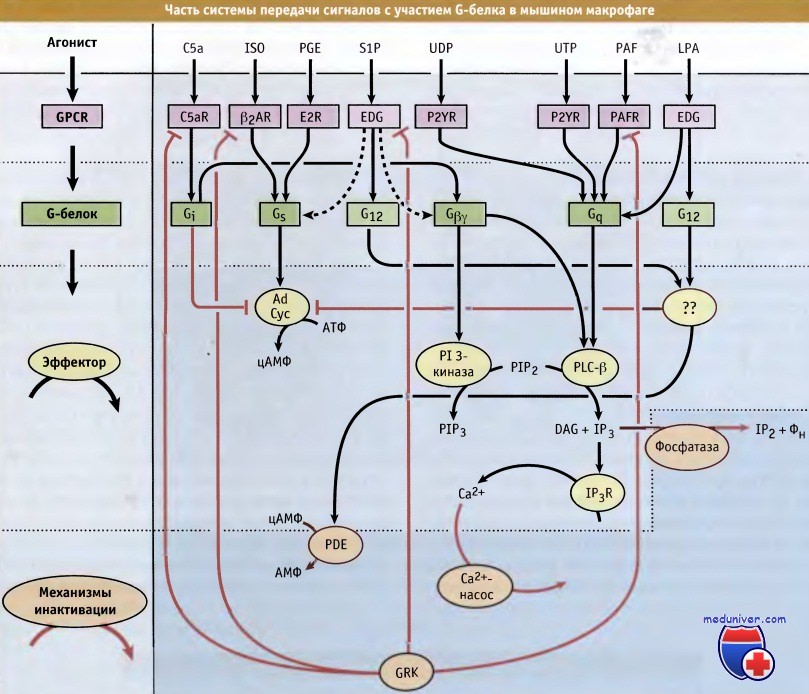
Участок сети передачи сигнала с участием G-белка у макрофага, который подчеркивает всю сложность взаимодействий, происходящих в такой системе.
Некоторые рецепторы и G-белок не показаны. Когда показан определенный G-белок, его выходной сигнал, вероятно, опосредуется его субъединицей Ga.
При активации любого G-белка также активируется его Gβγ-субъединица, хотя передача сигнала с участием этой субъединицы обычно наиболее выражена для Gi-тримеров.
Наряду с этим, некоторые G-белки модулируют активность других, пока еще недостаточно исследованных сигнальных путей.
Показано лишь немного эффекторов и единственный адаптивный механизм, представляющий собой фосфорилирование рецепторов, катализируемое GRK.
Данные предоставлены Полом Стернвайсом, Группа изучения передачи сигналов.
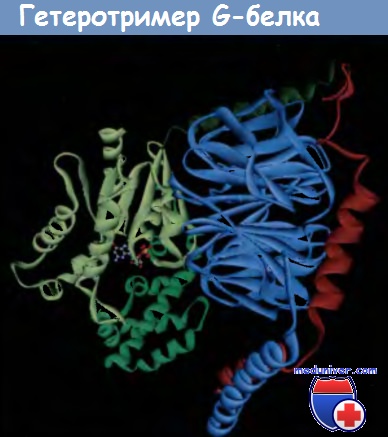
Структура неактивированного Gi-гетеротримера.
Каждая субъединица G-белка, ответственного за ингибирование аденилатциклазы и за передачу сигналов с участием Gβγ, выделена соответствующим цветом.
Показана связь GDP с субъединицей Gai. Структура построена по данным protein Data Bank file 1GP2.
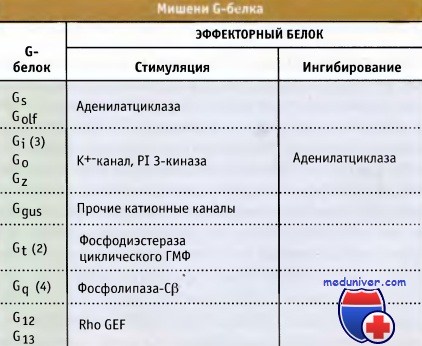
Эффекторы, находящиеся под контролем G-белка, не обнаруживают сходства структуры.
Они могут представлять собой ионные каналы или интегрированные в плазматическую мембрану ферменты,
белки, расположенные на внутренней стороне мембраны, или растворимые белки, способные связываться с субъединицами Ga.
В таблице представлены G-белки, подразделяющиеся на группы в соответствии со степенью гомологии их первичной структуры,
а также некоторые эффекторы, которые они контролируют.