Ферменты рестрикции. Применение в молекулярной генетике
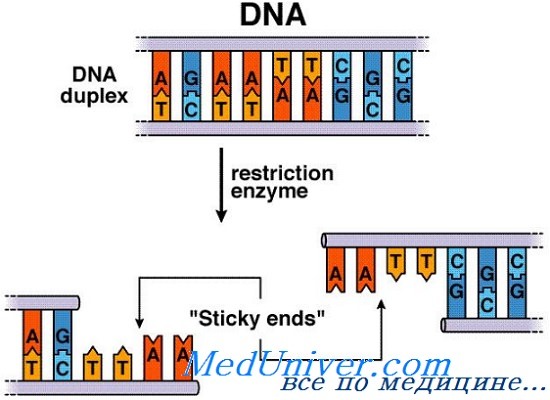
Прорывом в разработке молекулярного клонирования стало открытие в начале 1970-х годов бактериальных эндонуклеаз (рестриктаз) — ферментов, которые распознают специфические последовательности в двойной спирали ДНК и расщепляют обе нити ДНК в месте (сайте) распознавания. Разрывы могут быть прямо противоположны друг к другу, в этом случае они оставляют законченные нити ДНК, или могут быть смещены на несколько пар оснований в любом направлении, что приводит к тому, что один из концов нити ДНК «нависает» над другим.
В настоящее время известно более 3500 ферментов рестрикции, каждый со своим собственным сайтом распознавания, состоящим из четырех или шести пар оснований, хотя несколько ферментов имеют более длинные сайты. Последовательность оснований сайта обычно бывает палиндромом, т.е. совпадает при чтении как с 5', так и с 3'-конца нити ДНК.
Например, фермент рестрикции ЕсоRI распознает специфическую палиндромную последовательность из шести пар оснований 5'-GAATTC-3, где бы они ни находились в двойной молекуле ДНК.
Фермент расщепляет ДНК в этом месте, со смещением разрывов в нитях ДНК в 4 пары оснований между гуанином и смежным аденином в последовательности GAATTC. При этом образуется два фрагмента, каждый с однонитевым окончанием из четырех оснований 5'-ААТТ-3' в конце.
Расщепление молекулы ДНК конкретным ферментом рестрикции разрывает ДНК на характерный и воспроизводимый набор фрагментов, распределение длин которых отражает частоту и позиции специфических мест воздействия фермента. Например, рестриктаза ЕсоRI разрывает двойную спираль ДНК специфично в последовательности 5'-GAATTC-3'.
Обработка ДНК всего генома человека ферментом ЕсоRI генерирует набор приблизительно из 1 млн фрагментов переменной длины, каждый с конкретной позицией в геноме. В среднем фермент с сайтом опознавания в 6 пар оснований, подобно ЕсоRI, должен расщеплять ДНК человека через каждые 46 пар оснований, или каждые 4096 пар оснований. В действительности такие места не распределены равномерно из-за состава оснований и их последовательности на протяжении генома.
Таким образом, ЕсоRI фрагментирует ДНК на куски размером от десятка до многих сотен тысяч пар оснований; длина каждого фрагмента определяется тем, сколько ДНК находится между двумя последовательными сайтами распознавания ЕсоRI.
Поскольку все молекулы ДНК, обработанные ферментом ЕсоRI, независимо от их происхождения, имеют идентичные однонитевые концы, любые две ДНК молекулы, обработанные ЕсоRI, могут быть соединены вместе in vitro, путем спаривания комплементарных концов, содержащих 4 пары оснований, с помощью фермента ДНК-лигазы. Это создает рекомбинантную молекулу ДНК, один конец которой происходит из одного источника, а второй — от другого.
Когда фермент рестрикции разрезает обе нити в одной и той же позиции, оставляя «тупые» концы, ДНК-лигаза также может соединять их вместе без необходимости в совместимости «нависающих» нитей.