• Во всех эукариотических клетках на пути экзоцитоза находятся одни и те же основные компартменты: эндоплазматический ретикулум, компартменты аппарата Гольджи и транспортные везикулы, осуществляющие дальнейший перенос от аппарата Гольджи
• Количество и строение органелл экзоцитоза варьируют от организма к организму и от клетки к клетке
• Каждая органелла на пути экзоцитоза обладает специализированной функцией
• Эндоплазматический ретикулум является местом синтеза и сворачивания белков
• В аппарате Гольджи происходит модификация, сортировка и дальнейший перенос белков к месту назначения с помощью транспортных везикул
• Транспорт карго к плазматической мембране происходит или непосредственно, за счет конститутивного процесса, или опосредованно, за счет регулируемого процесса, который включает временное хранение белков в секреторных гранулах до момента получения клеткой соответствующего сигнала
Экзоцитоз представляет собой один из путей перемещения белков, предназначенных для плазматической мембраны или на экспорт во внеклеточное пространство. Ранее этот путь назывался секреторным, поскольку впервые был обнаружен в специализированных клетках, названных панкреатическими ацинарными клетками, которые секретируют пищеварительные ферменты. Ферменты составляют большую часть секретируемых белков. Этот факт был использован Паладом с сотрудниками для того, чтобы проследить маршрут этих белков.
Ими были проведены пульс-чейз эксперименты, в которых срезы ткани поджелудочной железы инкубировали с радиоактивно меченными аминокислотами, и в течение определенного времени регистрировали положение радиоактивных белков методом радиоавтографии. Исследователи показали, что пищеварительные ферменты синтезируются в ЭПР и транспортируются оттуда в аппарат Гольджи.
Затем ферменты обнаруживаются в уплотняющихся гранулах, которые после созревания становятся зимогеновыми гранулами. Эти гранулы локализованы поблизости от участка плазматической мембраны, выстилающего проток, по которому ферменты поступают в тонкий кишечник. Высвобождение ферментов происходит только в процессе регулируемой секреции, когда клетка получает соответствующий стимул, например воздействие гормона, который высвобождается в пищеварительный тракт под действием пищи.
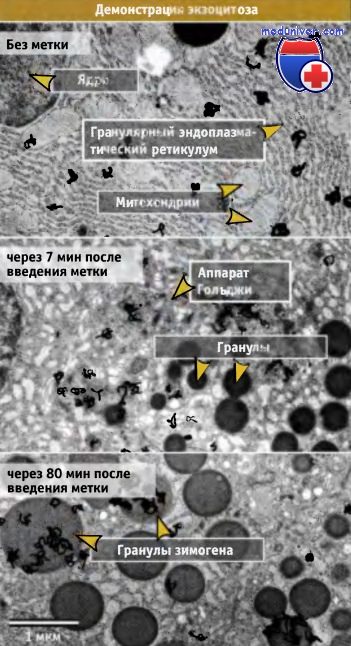
Механизм секреторного транспорта был выяснен при исследовании биогенеза гранул зимогена.
Срезы поджелудочной железы импульсно метили Н3 лейцином, инкубировали в течение 0, 7 и 80 мин и фиксировали.
После фиксации готовили препараты для электронной микрорадиографии.
Темные зерна, соответствующие вновь синтезированным секреторным белкам, вначале локализуются над ЭПР (верхний снимок),
затем над аппаратом Гольджи (средний снимок), и пото'м над секреторными/зимогеновыми гранулами (нижний снимок).
В отличие от регулируемой, конститутивная секреция представляет собой процесс, при котором секреция белков происходит непрерывно. Эта форма преобладает в клетках, для которых нехарактерна регулируемая секреция. Например, клетки печени постоянно секретируют белки в плазму крови (альбумин). При регулируемой и конститутивной секреции содержащие белок везикулы сливаются с плазматической мембраной в результате процесса, называемого экзоцитоз.
Организация пути экзоцитоза в ацинарных клетках поджелудочной железы показана на рисунке ниже. ЭПР представлен единственным компартментом с четко выраженными доменами, среди которых наиболее важную роль в секреции играет гранулярный, или шероховатый, ЭПР (ГРЭПР). «Шероховатость» его обусловлена связанными рибосомами, синтезирующими секреторные белки, которые переносятся через липидный бислой в люмен ЭПР. Уплощенные цистерны ЭПР упакованы вместе и занимают базальную часть клетки.
Большое количество элементов эндоплазматического ретикулума (ЭПР) в клетках поджелудочной железы обеспечивает достаточное количество рибосомальных сайтов связывания и транслокации для 10 млн пищеварительных ферментов, которые синтезируются в них каждую минуту. В большинстве клеток эукариот мембраны ЭПР представляют собой наиболее распространенные внутриклеточные мембраны. Даже в клетках, не специализированных на секрецию белков, на мембраны ЭПР обычно приходится около 50% от всех клеточных мембран.
Вновь синтезированные секреторные белки переносятся от эндоплазматического ретикулума (ЭПР) в аппарат Гольджи, который представляет собой органеллу, содержащую множество ком-партментов. Характерной особенностью органеллы является наличие стопок Гольджи, или диктиосом, представляющих собой вертикальные ряды уплощенных цистерн, напоминающих стопки пита хлеба (плоский круглый хлеб с кармашком внутри, который можно заполнить любой начинкой). Стопка Гольджи обладает полярностью и состоит из цис-, медиа- и транс-цистерн.
Цис-цистерна расположена со стороны входа в аппарат Гольджи, а транс-цистерна находится у выхода из нее. Расширенные края цистерн представляют собой места, откуда отпочковываются транспортные везикулы, и с которыми происходит их слияние.
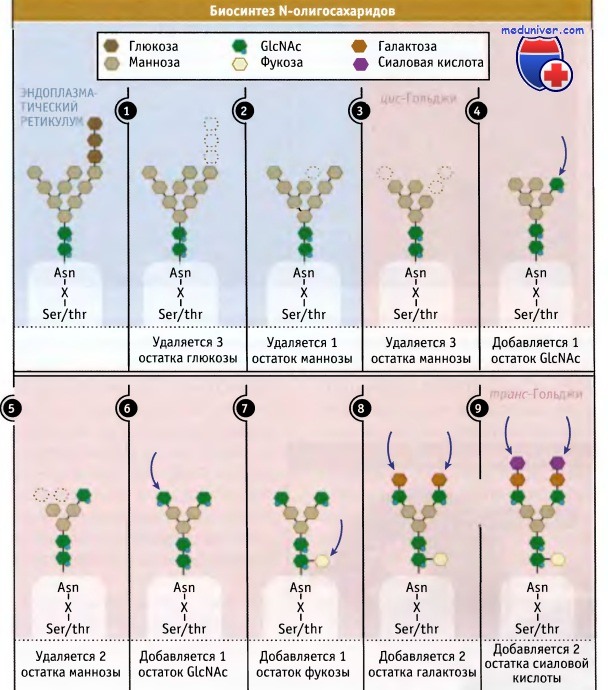
Цистерны Гольджи содержат различные ферменты, которые осуществляют посттрансляционные модификации большинства транспортируемых белков. Лучше всего охарактеризованы ферменты, которые модифицируют О- и N-олигосахариды, связанные с белками. Эти олигосахариды выполняют много функций, включая адресование в лизосомы вновь синтезированных лизо-сомальных ферментов.
Как показано на рисунке ниже для N-олигосахаридов, модификация остатков, содержащих много маннозы и присоединенных к ЭПР, происходит ступенчатым образом и приводит к постепенному превращению «незрелых» олигосахаридов, содержащих много маннозы, в сложные, богатые сиаловыми кислотами структуры Считается, что ферменты, ответственные за эти этапы, расположены друг за другом вдоль стопки цистерн. Ферменты, принимающие участие на ранних этапах этого процесса, вероятно, находятся на входящей стороне аппарата Гольджи, которая первая получает белки, экспортированные из ЭПР.
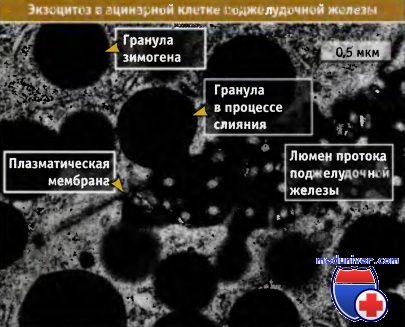
Фотография в трансмиссионном электронном микроскопе,
показывающая гранулы зимогена, сливающиеся с плазматической мембраной.
Содержимое гранул высвобождается в проток поджелудочной железы.
Ферменты, которые действуют позже (например, осуществляющие добавление остатков сиаловых кислот), обнаруживаются на стороне выхода, откуда секретируемые белки покидают аппарат Гольджи.
Стопки Гольджи с каждой стороны ограничены ретикулотубулярной сетью. Цис-Гольджи сеть (CGN), расположенная с входной стороны аппарата Гольджи, получает белки, выходящие из экспортных сайтов ЭПР. Она осуществляет функции контроля качества и возвращает на место резидентные белки, которые вышли из ЭПР. Транс-Гольджи сеть (TGN), со стороны выхода, распределяет белки по разным направлениям. В ацинарных клетках поджелудочной железы большинство белков в TGN представляют собой секреторные белки конденсирующихся вакуолей, которые созревают в зимо-геновые гранулы.
Также в TGN лизосомальные белки отделяются от белков плазматической мембраны и постоянно секретируемых белков. Первые транспортируются в лизо-сомы (по эндосомальному пути), а вторые направляются к клеточной поверхности. В клетках, не осуществляющих функцию регулируемой секреции, образуются только лизосомальные белки и белки плазматической мембраны.
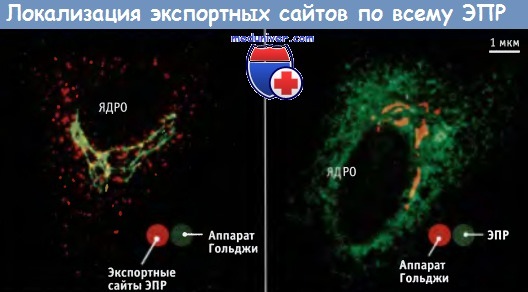
Хотя для аппарата Гольджи характерна цис-, транс-полярность, отражающая направление транспорта, в ЭПР она отсутствует. Вновь синтезированные секреторные белки направляются к экспортным сайтам ЭПР, т. е. к регионам, в которых происходит образование транспортных везикул. В клетках млекопитающих эти экспортные сайты локализованы по всему ГРЭПР и не всегда связаны с аппаратом Гольджи. В клетках, в которых регулируемая секреция отсутствует, экспортные сайты ЭПР часто расположены на значительном расстоянии от аппарата Гольджи. Как показано на рисунке ниже, сеть ЭПР распространена по всей цитоплазме, и в ней случайным образом располагаются порядка сотни экспортных сайтов.
Напротив, аппарат Гольджи обычно расположен поблизости от ядра. Поэтому некоторые везикулы, переносящие секреторные белки от ЭПР, прежде чем достигнуть аппарата Гольджи, должны преодолеть расстояние в несколько микрон.
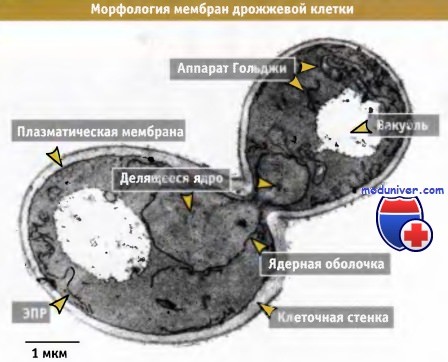
Так же как ацинарные клетки поджелудочной железы являются идеальным объектом для исследования роли секретируемых белков методами микроскопии и биохимического фракционирования, и почкующиеся дрожжи Saccbaromyces cerevisiae оказались подходящей генетической системой для изучения молекулярных механизмов перемещения белков. Как следует из рисунка ниже, почкующиеся дрожжи обладают набором органелл, необходимых для экзоцитоза и по функциям, напоминающим соответствующие структуры клеток млекопитающих. Однако по организации они несколько отличаются.
В клетках дрожжей эндоплазматического ретикулума (ЭПР) выглядит менее плотным и в основном локализован непосредственно у плазматической мембраны и вокруг ядра, ЭПР не содержит явных экспортных сайтов, и транспортные везикулы, вероятно, образуются из любых мест мембраны ЭПР. Аппарат Гольджи не формирует стопок цистерн и не сосредоточен у ядра. У дрожжей отсутствуют секреторные гранулы, и для них нехарактерен регулируемый экзоцитоз. Несмотря на эти различия, основной транспортный механизм у дрожжей гомологичен соответствующему механизму в клетках животных и является достаточно консервативным в клетках эукариот.
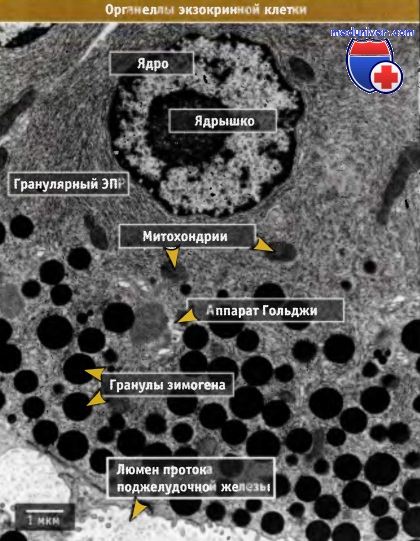
Фотография в трансмиссионном электронном микроскопе, показывающая органеллы в ацинарной клетке поджелудочной железы.
В экзоцитозе участвует гранулярный эндоплазматический ретикулум, аппарат Гольджи и гранулы зимогена.
В этих клетках особенно развит гранулярный ЭПР и много гранул зимогена.
Этапы образования N-олигосахаридов;
везикулярный транспорт гликопротеинов между компартментами не показан.
Локализация белков в эндоплазматическом ретикулуме,
аппарате Гольджи и в экспортных сайтах ЭПР по данным иммунофлуоресцентной микроскопии.
Показаны две разных клетки.
Фотография в трансмиссионном электронном микроскопе,
показывающая почкующуюся дрожжевую клетку.