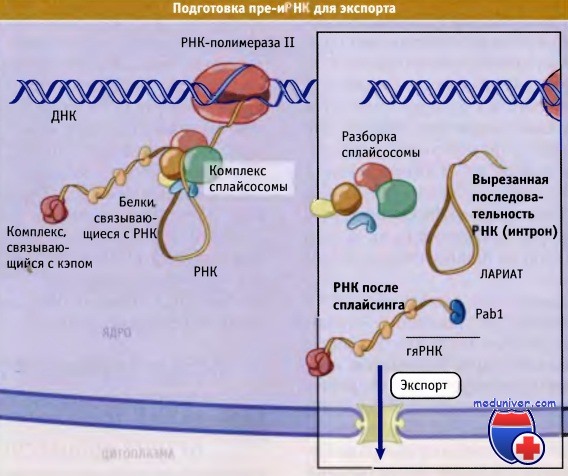
• Белки, которые связываются с иРНК в процессе транскрипции, отмечают места процессинга на пре-иРНК, и, вероятно, также участвуют в упаковке иРНК для экспорта
• Большинство белков в ядре, связанных с иРНК, после экспорта теряют с ней связь и возвращаются в ядро. Некоторые белки удаляются непосредственно перед экспортом
• На белках, связанных с иРНК, могут присутствовать сигналы для ее экспорта
• Экспорт иРНК может регулироваться, однако механизм этой регуляции остается неизвестным
Как уже отмечалось, большинство РНК образуются в ядре и экспортируются в цитоплазму, где осуществляют свои функции. Все процессы биогенеза иРНК, происходящие в ядре, начиная от транскрипции и заканчивая экспортом, являются взаимосвязанными. РНК не экспортируются в свободном виде, а в комплексе с белками.
В случае иРНК, с ее предшественниками (пре-иРНК) по мере синтеза ассоциируются многочисленные белки. Образующиеся РНК-белковые комплексы называются гетерогенные ядерные рибонуклеопротеиновые частицы (гяРНП частицы). Соответствующие белки (отличные от кэпсвязывающих и поли(А) связывающих белков) называются белками гя-РНП частиц.
В клетках человека присутствует по крайней мере 20 различных белков гяРНП частиц. Некоторые из них участвуют в формировании структуры пре-иРНК, необходимой для того, чтобы процессинг осуществлялся в нужном направлении. Это важно, поскольку экспортируются только те иРНК, которые прошли полный и правильный процессинг.
Используя инструменты, позволяющие отличать иРНК, которые прошли неправильный процессинг, клетка предотвращает экспорт дефектных иРНК, способных образовывать неправильные белки, присутствие которых может оказаться для нее вредным.
Считается, что функция многих белков, связывающихся с иРНК в ядре, состоит в облегчении ими упаковки иРНК, предназначенной для экспорта. Большинство клеток эукариот содержит от 3000 до 15 000 различных иРНК. Эти РНК сильно варьируют по размерам, последовательности и вторичной структуре, которая образуется при скручивании их молекул.
Предполагают, что, образуя РНК-белковые комплексы до начала экспорта, различные иРНК приобретают общие структурные особенности, важные для их эффективного экспорта. Некоторые из белков, связанных с иРНК, могут содержать специфические сигналы экспорта.
Ассоциация белков с иРНК носит временный характер. Некоторые из них удаляются до момента прохождения гяРНП частицы через ЯПК. Другие сопровождают РНК во время выхода из ядра. После транслокации через ЯПК, большая часть остающихся белков удаляется и возвращается обратно в ядро для участия в экспорте других иРНК.
гяРНП ассоциируют с иРНК в момент ее синтеза.
После сплайсинга сплайсосома и вырезанный лариат остаются в ядре,
а иРНК со связанными белками гяРНП из него выходит.
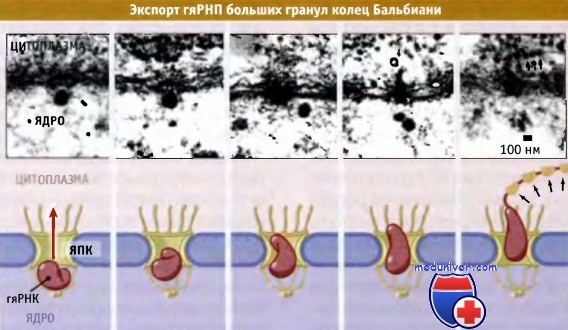
Поскольку гяРНП представляют собой большие комплексы, поскольку для того, чтобы пройти через ЯПК, они могут изменить свою конформацию. Уникальная особенность клеток насекомого Chironomus tentans позволяет наблюдать стадии экспорта гяРНП из ядра. На определенной стадии развития, когда происходит индукция транскрипции нескольких генов, этот организм образует набор крайне больших иРНК.
Сайты, на которых происходит активный процесс транскрипции, называются кольцами Бальбиани. Образующиеся иРНК характеризуются очень большими размерами, и, подобно другим видам иРНК, связаны с гяРНП частицами. Эти частицы называются гранулами колец Бальбиани. Как показано на рисунке ниже, они транспортируются через ядерные поры.
Диаметр гранул кольца Бальбиани гораздо больше диаметра канала ЯПК, который составляет около 27 нм. Гранулы можно визуализировать в электронном микроскопе, исследуя клеточные препараты, зафиксированные на разных стадиях процесса транспорта Как показано на рисунке ниже, при прохождении через ЯПК гранулы колец Бальбиани принимают линейную форму.
Линейная форма гяРНП частицы настолько длинна, что она заполняет ЯПК, и концы ее выдаются с обеих сторон поры. Поскольку вдоль гранулы кольца Бальбиани существуют структурные различия, мы знаем, что они всегда проходят через ЯПК в одинаковой ориентации, при которой ведущим является 5'-конец иРНК. Мы не знаем, каким образом экспортируются другие иРНК, но полагаем, что при необходимости их конформация может меняться.
Различные иРНК, которые образуются в клетке, обладают лишь немногими общими чертами, способными функционировать в качестве сигналов экспорта. Однако такие черты могут быть свойственны белкам, и вполне вероятно, что один или несколько белков гяРНП частиц содержат сигнал ядерного экспорта. Согласно этому представлению, белки, обладающие NES, связываются с иРНК, а транспортный рецептор узнает NES. По крайней мере на некоторых белках гяРНП частиц находятся NES, а на других белках они отсутствуют.
Экспорт некоторых иРНК происходит с участием сигналов, которые локализованы на белках, связывающихся с иРНК, но не принадлежащих к белкам гяРНП частиц.
Хотя обычно до завершения процессинга иРНК не экспортируются, известно, что некоторые РНК вирусов, содержащие интронные последовательности, могут выходить в цитоплазму. Ярким примером тому является РНК ВИЧ-1. Как и другие ретровирусы, ВИЧ-1 продуцирует набор перекрывающихся молекул иРНК, и некоторые из них подвергаются сплайсингу. Однако экспортироваться также должны полноразмерные вирусные РНК, не прошедшие сплайсинг, поскольку они представляют собой форму вирусной РНК, необходимую для воспроизводства новых инфекционных вирусных частиц. Также они функционируют как иРНК при образовании некоторых ВИЧ-белков.
Для того чтобы обеспечить экспорт иРНК, не прошедших сплайсинг, ВИЧ-1 обладает специальным механизмом. При трансляции одного из вариантов сплайсинга иРНК вируса образуется белок Rev. Этот белок связывается с последовательностью, которая присутствует в иРНК ВИЧ-1, не прошедших сплайсинг, и называется участок активации Rev (RRE). Белок Rev содержит NES, богатый лейцином. Этот сигнал участвует в экспорте как свободного белка Rev, так и связанного с иРНК, которая содержит RRE.
Сигнала RRE оказывается достаточным для того, чтобы обеспечить связывание Rev и экспорт иРНК в цитоплазму. При этом экспорт происходит таким же образом, как и экспорт белка, с участием экспортина 1.
Как и экспорт белка, экспорт иРНК меняется в ответ на внешний стимул и изменения внутриклеточной среды. Наилучшим примером регуляции экспорта иРНК в соответствии с реакцией клетки служит такое стрессорное воздействие, как температура. Эта реакция носит название тепловой шок, хотя она также развивается и при других видах стресса (например, при осмотическом шоке, действии токсических металлов или этанола в высокой концентрации).
Реакция теплового шока проявляется также в том, что большинство полиаденилированных РНК не экспортируются и накапливаются в ядре. В клетке происходит экспрессия генов теплового шока, которые кодируют белки, играющие защитную роль, и из ядра должны выходить иРНК белков теплового шока. Поэтому при тепловом шоке предотвращается экспорт некоторых иРНК и происходит экспорт иРНК белков теплового шока. Механизм такого переключения экспорта остается невыясненным.
Мы знаем, что регуляция экспорта иРНК при тепловом шоке не требует синтеза новых белков и может быть обнаружена уже через несколько минут после наступления теплового шока. Это позволяет предполагать, что индуцируется система передачи сигнала, изменяющая аппарат ядерного транспорта таким образом, что он начинает селективно экспортировать иРНК стрессорных белков или биогенез иРНК меняется в таком направлении, что правильно упаковываются и идентифицируются для экспорта только те иРНК, которые обеспечивают синтез стрессорных белков.
Экспорт гяРНП больших гранул колец Бальбиани С. tentans, наблюдаемый с помощью электронного микроскопа.
Схематические изображения иллюстируют различные стадии транспорта, наблюдаемые на микрофотографиях.
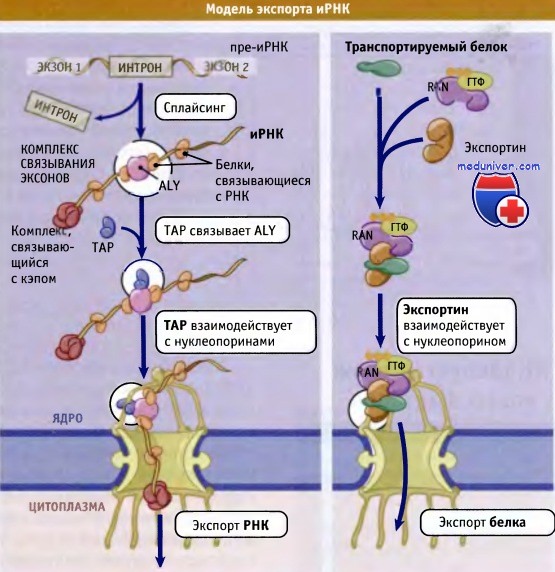
ТАР связывается как с иРНП, так и с ЯПК и функционирует как экспортный рецептор для иРНК.
Функция ТАР аналогична функции экспортина при транспортировке белков
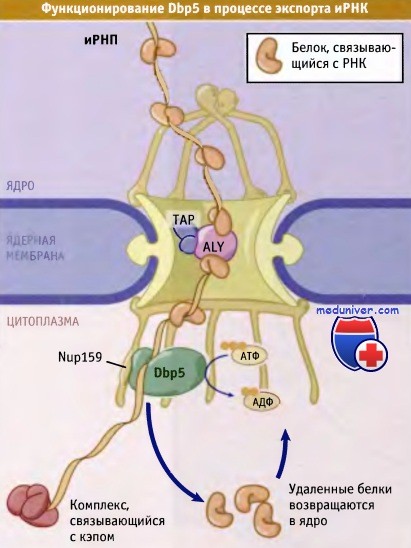
Dbp5 взаимодействует с терминальными филаментами ЯПК,
где он использует энергию АТФ для того, чтобы в процессе экспорта иРНК удалить белки иРНП.