• Щелевые контакты являются белковыми структурами, обеспечивающими прямой перенос небольших молекул между смежными клетками. Они присутствуют в большинстве клеток животных
• Щелевые контакты состоят из группы белковых каналов цилиндрической формы, направленных от плазматической мембраны и располагающихся между смежными клетками, в промежутке 2-3 нм
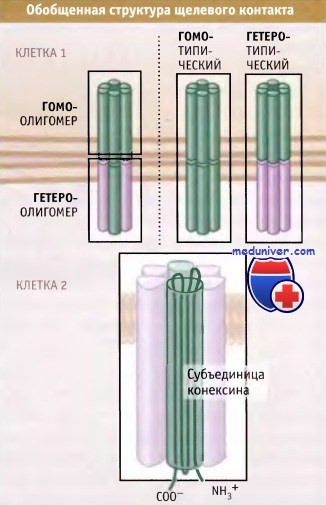
• Каналы щелевых контактов состоят из двух половин, которые называются коннексоны или полуканалы.
Каждый коннексон содержит шесть белковых субъединиц, называемых коннексинами
• У человека обнаружено более 20 разных коннексиновых генов. Все разнообразие типов коннексонов обеспечивается сочетаниями этих генов
• Щелевые контакты допускают свободную диффузию молекул массой 1200 дальтон и исключают молекулы, масса которых достигает 2000 дальтон
• Проницаемость щелевых контактов регулируется открытием и закрытием ворот канала. Этот процесс носит название «гейтинг» (воротный механизм). Он контролируется изменением внутриклеточного pH, потоком ионов кальция или прямым фосфорилированием субъединиц коннексина
• Обнаружены еще два семейства белков щелевых контактов, не относящихся к коннексинам. Это позволяет предполагать, что в ходе эволюции животных происходило неоднократное возникновение щелевых контактов
Щелевые контакты представляют собой специализированные структуры клеточной поверхности, которые обеспечивают прямой перенос ионов и небольших молекул между смежными клетками. Они присутствуют в большинстве типов клеток позвоночных и беспозвоночных и представляют собой единственную из известных структур, обеспечивающую межклеточный транспорт у животных. (В клетках растений эти функции выполняют плазмодесмы.)
Щелевые контакты, существующие между клетками миокарда, также обеспечивают проведение электрического импульса, необходимое для сокращения сердечной мышцы.
Существование щелевых контактов было продемонстрировано в 1960-е годы, когда исследователи обнаружили, что электрический импульс проходит между смежными клетками непосредственно, а не через разделяющий их слой жидкой среды. Это позволило предположить, что клетки обмениваются заряженными ионами и другими небольшими молекулами через каналы, которые проходят через плазматическую мембрану и прямо соединяют смежные клетки.
Это предположение подтвердилось микрофотографиями, сделанными в электронном микроскопе, на которых были видны узкие «промежутки» в 2-3 нм, разделяющие плазматические мембраны рядом расположенных клеток.
Как видно на рисунке ниже, через эти промежутки проходят каналы щелевых контактов, которые расположены отдельными кластерами (или бляшками). При исследовании в электронном микроскопе препаратов, полученных методом замораживания-скалывания, видно, что каналы расходятся от плазматических мембран клеток. Щелевые контакты могут содержать от нескольких десятков до многих тысяч каналов, и на клеточной поверхности их диаметр в сумме может достигать нескольких мкм.
Основным структурным элементом щелевого контакта является коннексон,
который состоит из шести трансмембранных коннексиновых субъединиц.
Длина каждого коннексона составляет 17 нм, а диаметр 7 нм.
Каналы состоят из двух половин, которые называются полуканалами, или коннексонами. Эти половины стыкуются в межклеточном промежутке. Каждый коннексон состоит из шести белковых субъединиц, которые называются коннексинами.
Коннексон представляет собой гидрофильный канал цилиндрической формы, длиной 17 и диаметром 7 нм в наиболее широкой части и около 3 нм в наиболее узкой. Центральные поры каналов видны в микроскопе при исследовании препаратов, приготовленных методом негативного контрастирования. Субъединицы коннексина содержат четыре трансмембранных альфа-спирали, связанные двумя внеклеточными петлями. Структура позволяет предполагать, что внеклеточные петли противоположных коннексинов связываются между собой посредством антипараллельных b-слоев, образуя b-цилиндр (b-бочонок).
Как следует из рисунке ниже, щелевые контакты обладают различным составом. Результаты, полученные при секвенировании генома человека, указывают, что в наших клетках существует, по меньшей мере, 20 различных коннексиновых белков. Во многих клетках экспрессируется более одного типа коннексинов. Это приводит к образованию гомо-олигомерных (состоящих из субъединиц только одного типа) и гетеро-олигомерных коннексонов (содержащих субъединицы разных типов).
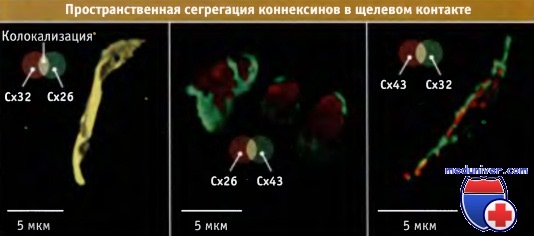
Вместе с тем, коннексоны могут стыковаться с одинаковыми (гомотипичными) или разными по составу (гетеротипичными) каналами. В одной бляшке щелевого контакта могут содержаться коннексоны, различные по составу коннексинов. Как представлено на рисунке ниже, в бляшке коннексоны или гомогенно расположены, или пространственно разобщены по коннексиновому составу.
Идентифицированы специальные домены, участвующие в стыковке коннексон/коннексон, узнавании коннексин/коннексин, образовании олигомеров и в сочетаемости (селективности) коннексиновых субъединиц. Существование этих доменов было продемонстрировано в экспериментах по связыванию рекомбинантных мутантных коннексинов, у которых отсутствовали специфические участки молекулы, или химерных коннексинов, состоящих из участков различных коннексинов. В настоящее время исследуется, каким образом функционируют эти домены.
Вначале, для проверки гипотезы о том, что клетки используют каналы для обмена небольшими молекулами, в клетки, растущие в культуре, вводили флуоресцирующие молекулы. С помощью микроскопа наблюдали диффузию этих молекул во времени. Эти эксперименты показали, что молекулы проходят между соседними клетками гораздо быстрее, чем этого можно было бы ожидать, если бы они проходили через липидные слои плазматической мембраны. Результаты подтвердили существование прямых каналов, связывающих цитозоль соседних клеток. В дальнейшем эти каналы были идентифицированы как щелевые контакты.
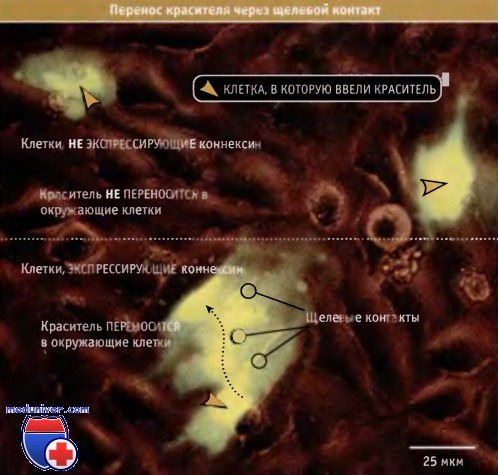
Используя флуоресцирующие молекулы различного размера, было показано, что через щелевой контакт могут проходить молекулы размером до 1200 дальтон (что соответствует диаметру около 2 нм), а молекулы более 2000 дальтон задерживаются. На рис. 15.59 представлены результаты недавно проведенного эксперимента по обмену флуоресцирующих молекул между клетками, экспрессирующими коннексины.
Подобные эксперименты демонстрируют, что ионы могут свободно обмениваться между цитозольными компартментами клеток, связанных щелевыми контактами. Клетки могут обмениваться и такими малыми молекулами, как сахара, нуклеотиды и вторичные мессенджеры (цАМФ и цГМФ). Межклеточные связи посредством щелевых каналов играют особенно критическую роль в тех случаях, когда большому количеству клеток необходимо генерировать быстрый, хорошо скоординированный ответ. Например, быстрые рефлекторные реакции в головном мозгу осуществляются с участием нейронов, связанных посредством щелевых контактов, которые обеспечивают практически моментальный обмен ионами; быстрый обмен ионов также обусловливает тщательно контролируемое во времени сокращение мышечных волокон миокарда.
Проницаемость щелевых контактов, вероятно, контролируется открытием и закрытием каналов (этот процесс называется гейтинг каналов, или воротный механизм). Существуют убедительные доказательства в пользу того, что гейтинг каналов находится под контролем протеинкиназ, которые фосфорилируют субъединицы коннексина, а также контролируется внутриклеточным pH и ионами кальция. Например, при увеличении концентрации ионов кальция с 10-7 М до 10-5 М проницаемость этих каналов падает; при концентрациях выше 10-5 М канал полностью закрывается.
Это, вероятно, служит механизмом клеточной самозащиты, поскольку при апоптозе происходит резкое возрастание внутриклеточной концентрации кальция, и закрытие щелевых каналов защищает соседние клетки, не позволяя им случайно включать механизм апоптоза.
Обнаружены еще два семейства белков щелевых контактов. Иннексины (коннексины беспозвоночных) найдены только у беспозвоночных и, несмотря на название, не являются гомологами коннексинов. Тем не менее они способны образовывать межклеточные контакты, которые внешне и по своим свойствам напоминают щелевые контакты позвоночных. Пользуясь «нексиновой» номенклатурой, следующее семейство называется паннексины (от лат. pan — весь, все). Паннексины обнаружены у позвоночных и у беспозвоночных и по структуре отличаются от коннексинов и иннексинов.
Паннексины почти исключительно находятся в нейронах, что позволяет предполагать, что они могут играть важную роль в развитии и функционировании этих клеток, даже у организмов с примитивной нервной системой. Описанные факты позволяют считать, что щелевые контакты возникли, по меньшей мере, дважды в ходе эволюции животных; это произошло совершенно независимым образом, в результате процесса, известного под названием конвергентной эволюции.
Субъединицы коннексина (Сх) в бляшках щелевого контакта.
Метод двойного иммунофлуоресцентного окрашивания. В клетки трансфецировали указанные пары коннексиновых генов и затем окрашивали их флуоресцентными антителами к соответствующим белкам.
Видно, например, что Сх32 локализуется вместе с Сх26, но не с Сх43.
Сами клетки невидимы.
Перенос красителя через щелевой контакт, наблюдаемый с помощью флуоресцентного микроскопа.
Проводили временную трансфекцию клеток ДНК, кодирующей коннексин, и, таким образом, не все клетки экспрессировали белок.