• Са2+-каналы, расположенные на поверхности клетки, переводят сигналы, получаемые мембраной, во внутриклеточные Са2+-сигналы
• Потенциал-зависимые Са2+-каналы представляют собой асимметричные белковые комплексы, состоящие из пяти различных субъединиц
• Подобно К+-каналу, а-субъединица потенциал-зависимых Са2+-каналов образует пору и содержит структуру поровой петли
• Селективный фильтр Са2+-канала образует электростатическую ловушку
• Са2+-канал стабилизируется в закрытом состоянии с помощью блокаторов канала
Ионы Са2+ представляют собой вторичный мессенджер и контролируют многие клеточные функции, такие как сокращение сердечной и скелетных мышц, зрительный процесс в сетчатке глаза, иммунный ответ Т-клеток, возбудимость нервных клеток и поведенческие реакции, а также секрецию инсулина b-клетками поджелудочной железы. Активация клеточных функций наступает в ответ на изменения концентрации Са2+ в цитозоле покоящейся клетки, которая примерно в 10 000 раз ниже, чем его концентрация во внеклеточной среде.
Запасы Са2+ в эндоплазматическом (ЭПР) и в саркоплазматическом ретикулуме (СР) достигают таких же величин, как его содержание во внеклеточной среде.
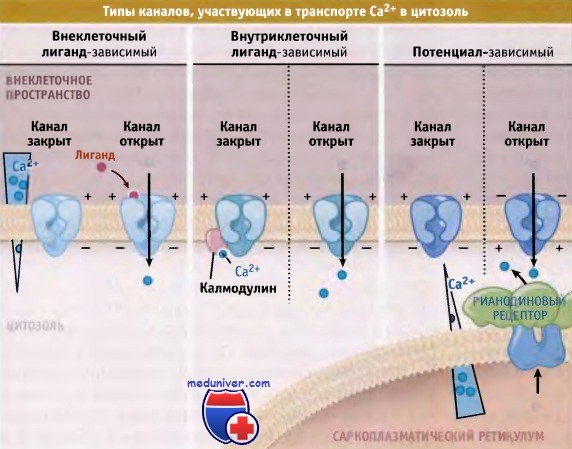
Уровень Са2+ в цитозоле регулируется за счет совместного функционирования различных растворимых Са2+ связывающих белков, а также белков, осуществляющих его трансмембранный перенос. Например, различные типы Са2+-каналов в плазматической мембране, в ЭПР и СР катализируют селективный транспорт ионов в цитозоль в направлении их электрохимического градиента. Различные типы каналов характеризуются различными воротными механизмами с участием внеклеточных лигандов, электрического потенциала, или же самих ионов Са2+.
При закрытии Са2+-каналов плазматической мембраны прерывается передача сигнала, и ионы Са2+ начинают выходить из цитозоля при участии специальных транспортных белков. Закрытие Са2+-каналов плазматической мембраны, которое предотвращает попадание в клетку избытка ионов Са2+, происходит с участием двух основных механизмов: Са2+-зависимой и потенциал-зави-симой инактивации. При активации белка калмодули-на, который связывается с внутриклеточным доменом Са2+-канала в ответ на поступление Са2+ в клетку, происходит инактивация многих типов Са2+-каналов по механизму отрицательной обратной связи.
В настоящем разделе мы рассмотрим предполагаемый механизм транспорта ионов через Са2+-каналы, основное внимание уделив потенциал-зависимым Са2+-каналам, и сравним механизмы их функционирования с работой К+-каналов.
Три основных типа каналов транспорта ионов Са2+ из внеклеточной среды в клетку.
Ионные каналы также осуществляют транспорт ионов Са2+ из внутриклеточных депо в цитозоль.
Слева показан градиент ионов Са+, создающийся по обеим сторонам плазматической мембраны и мембраны саркоплазматического ретикулума.
Потенциал-зависимые Са2+-каналы обеспечивают транспорт Са2+ в клетки в тех случаях, когда значения мембранного потенциала изменяются в положительную сторону за счет деполяризации. Эти каналы переводят электрический сигнал плазматической мембраны во внутриклеточный сигнал. Поступление в клетку ионов Са2+ приводит к увеличению его концентрации до определенного уровня и тем самым к запуску различных процессов, таких, как, например, мышечное сокращение, высвобождение гормонов и нейромедиаторов, активация Са2+-зависимых каскадных процессов и транскрипция генов. Существуют различные типы потенциалзависимых Са2+-каналов, которые классифицируются в соответствии с их электрофизиологическими и фармакологическими свойствами.
В данной статье мы рассмотрим Са2+-каналы L-типа, которые были клонированы первыми и остаются наиболее изученными. Каналы этого типа находятся в плазматической мембране скелетных мышц, миокарда и гладких мышц, а также нейронов. Активация их происходит при деполяризации мембраны.
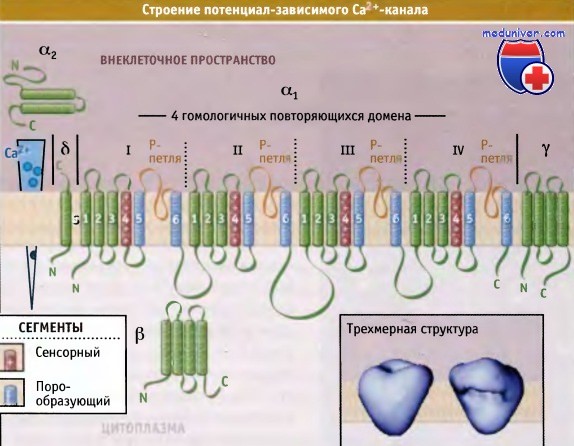
Эти каналы называются каналами L-типа потому, что они в течение долгого времени могут оставаться в открытом состоянии. В клетках скелетных и гладких мышц, миокарда, в нейронах, эндокринных клетках и сетчатке они представлены основными Cavl.X изоформами. Са2+-каналы L-типа представляют собой гетероолигомерные белковые комплексы, состоящие из пяти субъединиц: а1, а2, δ, b и γ. Функционирование канала и его позиционирование на мембране требуют участия всех субъединиц.
Присутствие гидрофобных и гидрофильных последовательностей в составе a1-субъединицы позволяет предполагать существование четырех порообразующих структур, каждая из которых состоит из трансмембранных сегментов 5 и 6 и из поровой петли, входящей в состав селективного фильтра. Также предполагается существование четырех сенсорных сегментов, каждый из которых состоит из трансмембранных сегментов 1-4. Такой тип трансмембранной организации напоминает потенциал-зависимые Na+-каналы. Субъединица b участвует в инактивации и закрытии канала. Вместе с а-субъединицей она может контролировать воротный механизм за счет взаимодействия с порообразующим трансмембранным сегментом S6 домена I.
При использовании метода электронной криомикроскопии были получены данные, позволившие предложить трехмерную структуру, описывающую строение потенциал-зависимого Са2+-канала L-типа. Как показано на рисунке ниже, канал представляет собой асимметричный белок, по форме напоминающий сердце и в широкой части снабженный подобием ручки. Большая часть локализована вне клетки и включает структуру наподобие ручки, а2-субъединицу, N-терминальный участок субъединицы δ и внеклеточные петли субъединиц а1 и у.
Предполагаемая структура субъединиц Са2+-канала L типа.
Внизу на вставке показана трехмерная структура,
полученная с использованием метода электронной криомикроскопии.
Са2+-каналы обладают высокой селективностью по отношению к ионам Са2+ и осуществляют транспорт ионов с высокой скоростью. Ионы Na+ относятся к числу самых распространенных внеклеточных катионов, и их концентрация почти в 100 раз превышает концентрацию ионов Са2+. Диаметр ионов Na+ близок к диаметру ионов Са2+ (2,0 А). Поэтому посредством простого молекулярного сита разделить их невозможно. Каким же образом Са2+-каналы достигают высокой селективности в отношении ионов Са2+ и в то же время обеспечивают их быстрый транспорт?
Поры К+-каналов обладают селективными фильтрами, которые имитируют водное окружение, стабилизирующее транспортируемые ионы. При прохождении через фильтр ионы подвергаются дегидратации. Например, селективные фильтры К+-каналов обладают четырьмя Р-петлями, образующими довольно жесткую структуру, изнутри облицованную атомами кислорода карбонильной группы. Этим достигается определенное расположение ионов К+ на пути транспорта. Селективный фильтр потенциалзависимых Са2+-каналов, вероятно, также образован четырьмя Р-петлями.
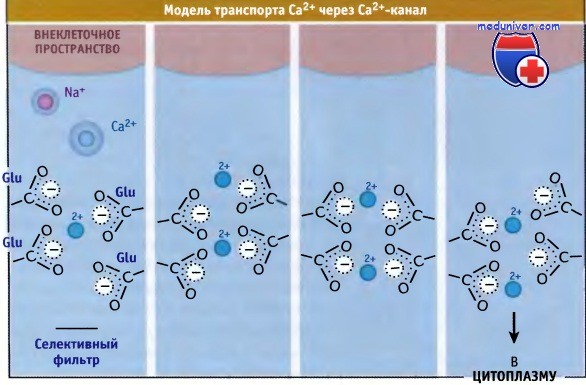
Однако полагают, что он представляет собой более гибкую структуру, каждая Р-петля которой содержит глутаминовый остаток, причем транспортный путь ионов облицован атомами кислорода не от карбонильных, а от карбоксильных групп. (Такая конструкция напоминает EF руку сайта связывания Са2+, когда ион располагается в полости, содержащей атомы кислорода, многие из которых относятся к карбоксильным группам.) Четыре глутаминовых остатка образуют т. н. локус ЕЕЕЕ, консервативную структуру, характерную для Са2+-каналов.
Локус ЕЕЕЕ контролирует предпочтительный транспорт ионов Са2+ по сравнению с другими катионами. Замещение любого из четырех остатков глутамата другими аминокислотами резко снижает селективность канала в отношении ионов Са2+. Предполагается, что специфическое связывание именно ионов Са2+ в области устья поры обеспечивается боковыми цепями глутаминовой кислоты. Это подтверждается данными in vitro электрофизиологических измерений, которые демонстрируют, что в отсутствие ионов Са2+ через Са2+-каналы начинают проходить ионы Na+. Ионы Na+ проходят через канал быстрее ионов Са2+. Более низкая скорость транспорта ионов Са2+ позволяет предполагать, что они прочнее связываются с порами канала. Это препятствует входу в пору ионов натрия, несмотря на то что во внеклеточном пространстве их содержится больше, чем ионов Са2+.
Сродство каналов к ионам Са2+ составляет около 10-6 М. Однако скорость транспорта, рассчитанная на основании этих данных, получается в 1000 раз ниже, чем измеренная, которая составляет 106 в 1 с. Для разрешения этого противоречия была предложена специальная модель. Согласно этой модели, локус ЕЕЕЕ способен связывать несколько ионов Са2+, и входящий в пору ион, благодаря возникающим силам электростатического отталкивания, вызывает высвобождение с внутренней стороны связанного иона Са2+. Таким образом, электростатическое отталкивание помогает преодолеть силу сродства связывания Са2+, которая бы замедлила его транспорт. Эта модель напоминает предложенную для К+-каналов, которые, как показано, обладают несколькими сайтами связывания катиона.
Потенциал-зависимые Са2+-каналы являются основными клиническими мишенями для лекарственных препаратов, используемых при лечении гипертонии и других заболеваний. К числу наиболее часто применяемых лекарственных средств относятся фенилалкиламины, бензотиазепины и дигидропиридины, которые обычно называют кальциевыми антагонистами. Первоначально, Са2+-антагонисты были введены в клиническую практику для лечении гипертонии как сосудорасширяющие средства. Они уменьшали уровень Са2+ в гладких мышцах, например в стенке кровеносных сосудов, приводя к снижению тонуса и кровяного давления.
Эксперименты по сайт-направленному мутагенезу и по связыванию лекарств позволили идентифицировать сайты, связывающие лечебные препараты. Они находятся в сегментах S5 и S6 III домена и в сегменте S6 IV домена, расположены с цитоплазматической стороны селективного фильтра. Ингибиторы ряда фенилалкиламина блокируют Са2+-каналы за счет прямого взаимодействия с остатками глутамата Р-петли селективного фильтра, которые находятся со стороны цитоплазмы, а дигидропиримидины и бензотиазепины проникают в пору канала со стороны наружного устья.
Гипотетический механизм, описывающий транспорт Са2+ через канал с участием ЕЕЕЕ-локуса.
Ионы Са2+ энергетически стабилизируют атомы кислорода карбоксильных групп и предотвращают транспорт ионов Na+,
которые не обладают достаточной величиной положительного заряда, чтобы попасть в локус ЕЕЕЕ и подвергнуться там эффективной дегидратации.
Силы электростатического отталкивания, существующие в поре между ионами Са2+, облегчают быструю диффузию.