• Гистоны ацетилируются в течение определенного времени при репликации
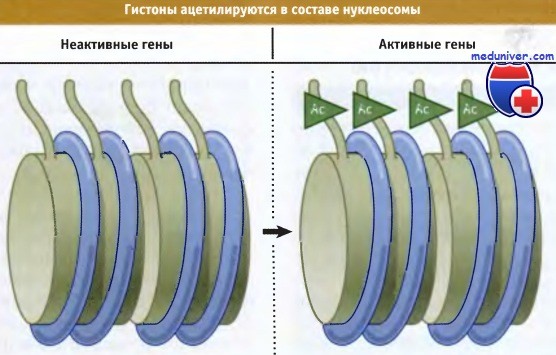
• Ацетилирование гистонов связано с активацией экспрессии генов
• Деацетилированный хроматин может обладать более конденсированной структурой
• Активаторы транскрипции связаны с ацетилазой гистонов в большие комплексы
• Комплекс ремоделирования хроматина может формировать комплекс ацетилирования
• Ацетилазы гистонов варьируют по специфичности к различным мишеням
• Ацетилирование может оказывать на транскрипцию качественное и количественное влияние
• Деацетилирование гистонов связано с репрессией активности генов
• Деацетилазы находятся в комплексе с репрессором генной активности
• Ацетилирование гистонов может являться событием, которое поддерживает комплекс в активированном состоянии
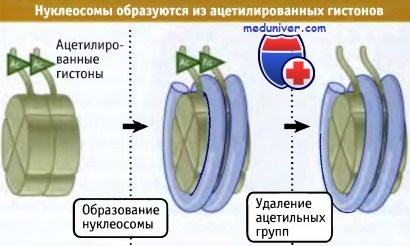
При репликации ацетилирование гистонов происходит до их включения в состав нуклеосом.
Все коровые гистоны могут подвергаться ацетилированию. Основные сайты ацетилирования представлены лизиновыми остатками, расположенными в N-терминальных участках гистонов Н3 и Н4. Ацетилирование происходит при протекании двух событий:
• репликации ДНК;
• активации генов.
При репликации хромосом в S-фазе клеточного цикла происходит ацетилирование гистонов. На рисунке ниже показано, что это ацетилирование происходит до включения гистонов в нуклеосомы. Известно, что гистоны Н3 и Н4 ацетилируются на стадии их объединения в тетрамер Н32-Н42. Затем тетрамер включается в нуклеосому. Вскоре после этого ацетильные группы удаляются.
Значение ацетилирования подчеркивается тем фактом, что подавление этой модификации гистонов Н3 и Н4 при репликации приводит к потере жизнеспособности клеток дрожжей. Два этих гистона представляются излишними в качестве субстратов, поскольку дрожжи в S-фазе узнают и могут ацетилировать любой из них. Ацетилирование гистонов может играть двоякую роль: оно может быть необходимо для узнавания гистонов факторами, включающими их в нуклеосомы, а также может потребоваться для реформирования существующих или сборки новых нуклеосом.
Ацетилированные и неацетилированные гистоны не различаются известными факторами, участвующими в сборке хроматина. Это позволяет предположить, что, скорее всего, модификации необходимы для последующих белковых взаимодействий. В течение долгого времени считали, что ацетилирование необходимо для осуществления контроля над белок-белковыми взаимодействиями, происходящими при объединении гистонов в нуклеосому. Доказательства в пользу такой роли ацетилирования были получены на дрожжах, у которых SAS гистон ацетилазный комплекс связывается с комплексами сборки хроматина на репликативной вилке, где он ацетилирует 16Lys в гистоне Н4. Это может являться частью системы, ответственной за определенный характер ацетилирования гистонов после репликации.
Вне S-фазы ацетилирование гистонов в хроматине обычно коррелирует с состоянием процесса генной экспрессии. Такая корреляция была установлена, поскольку ацетилирование гистонов увеличивается в доменах, содержащих активные гены, и ацетилированный хроматин более чувствителен к ДНКазе I и, возможно, к микрококковой нуклеазе. На рисунке ниже показано, что при этом происходит ацетилирование хвостов гистонов в нуклеосомах. Сейчас известно, что это происходит при активации гена, главным образом, из-за ацетилирования нуклеосом вблизи промотора.
Наряду с событиями, происходящими на индивидуальных промоторах, широкомасштабные изменения ацетилирования происходят в половых хромосомах. Это является частью механизма, посредством которого изменяется активность генов на Х-хромосоме с тем, чтобы скомпенсировать присутствие или двух Х-хромосом у одних организмов, или наоборот, только одной хромосомы (наряду с Y-хромосомой) у других. Неактивная Х-хромосома самок млекопитающих имеет недоацетилированный гистон Н4. Высокоактивная Х-хромосома у самцов Drosophila характеризуется повышенной степенью ацетилирования этого гистона. Это предполагает, что присутствие ацетильных групп может быть необходимо для создания более релаксированной активной структуры. У самца Drosophila происходит ацетилирование гистона Н4 Х-хромосомы по остатку 16Lys.
Компонент, ответственный за этот процесс, представляет собой фермент, называемый MOF, который связывается с хромосомой и представляет собой часть большого белкового комплекса. Этот комплекс «компенсации дозы» отвечает за общие изменения в Х-хромосоме, которые приводят к более высокому уровню экспрессии содержащихся в ней генов. Увеличение степени ацетилирования представляет собой проявление лишь одной из активностей, которыми обладает этот комплекс.
Ацетилирование, связанное с активацией генов,
происходит за счет прямой модификации гистонов в составе нуклеосомы.
Ацетилирование гистонов представляет собой обратимый процесс. Каждое направление реакции катализируется специфическим ферментом. Ферменты, способные ацетилировать гистоны, называются гистоновые ацетилтрансферазы, или HAT; удаление ацетильных групп катализируется деацетилазами гистонов, или HDAC. Известны две группы ферментов HAT: ферменты группы А действуют на гистоны в хроматине и контролируют транскрипцию; ферменты группы В действуют на вновь синтезированные гистоны в цитозоле и участвуют в сборке нуклеосом.
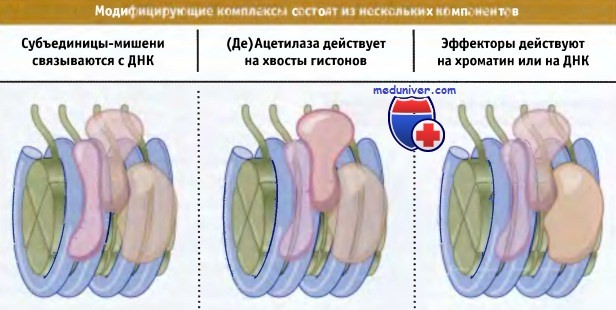
Общая черта ферментов ацетилирования состоит в том, что HAT группы А являются частью большого белкового комплекса. На рисунке ниже представлена упрощенная модель их функционирования. Обычно комплекс содержит субъединицы, которые связываются с определенными сайтами ДНК. Это определяет мишень для HAT. В состав комплекса также входят эффекторные субъединицы, способные изменять структуру хроматина или непосредственно влиять на транскрипцию. Вероятно, для того чтобы проявить свое действие, по крайней мере некоторые эффекторы требуют ацетилирования. Аналогичным образом может влиять деацетилирование, катализируемое HDAC.
Носит ли эффект ацетилирования количественную или качественную природу? Одна возможность заключается в том, что для проявления эффекта необходимо определенное количество ацетильных групп, а точное их расположение большей частью не играет роли. Альтернатива состоит в том, что отдельные события ацетилирования характеризуются специфическими эффектами. Мы можем объяснить существование комплексов, содержащих множественные активности HAT, следующим предположением — если индивидуальные ферменты проявляют различную специфичность, то множественная активность может понадобиться для ацетилирования множественных сайтов или для обеспечения различных посттранскрипционных эффектов. По-видимому, при репликации, по крайней мере в отношении гистона Н4, кажется, что ацетилирование по любым двум из трех доступных позиций является в этом случае адекватным. Это свидетельствует в пользу количественной модели. В тех случаях, когда меняется структура хроматина, что влияет на транскрипцию, важным становится ацетилирование в специфических сайтах.
Так же как ацетилазы вызывают активацию транскрипции, ее инактивация связана с функционированием деацетилаз. Это положение справедливо как для индивидуальных генов, так и для гетерохроматина. Репрессия индивидуальных промоторов достигается при образовании комплексов, обладающих деацетилазной активностью по отношению к отдельным участкам, находящимся вблизи промотора. Отсутствие ацетилированных гистонов в гетерохроматине характерно как для конститутивного (обычно расположенного в центромерной и теломерной области), так и факультативного гетерохроматина. Последний тип гетерохроматина в одних клетках находится в неактивном состоянии, а в других может быть активен. Обычно в гетерохроматине N-терминальные участки гистонов Н3 и Н4 не подвергаются ацетилированию.
Каким образом происходит доставка ацетилаз (или деацетилаз) к специфическим мишеням? Так же как это имеет место для комплексов ремоделирования хроматина, процесс, вероятно, осуществляется непрямым путем. Активатор (или репрессор), специфически узнающий определенную последовательность, взаимодействует с компонентом ацетилазы (или деацетилазы) комплекса, обеспечивая его позиционирование на промоторе.
Комплексы, участвующие в ремоделировании хроматина и в модифицировании гистонов, могут непосредственно взаимодействовать. Связывание SWI/SNF комплекса ремоделирования может повлечь за собой связывание ацетилазного комплекса. Ацетилирование гистонов может стабилизировать связь с комплексом SWI/SNF, тем самым взаимно способствуя функциям этих компонентов.
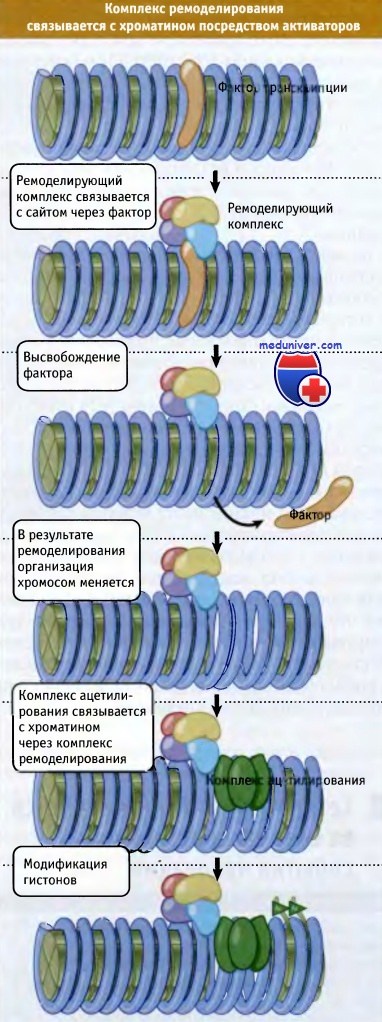
Серия событий, происходящих в области промотора, представлена на рисунке ниже. Вначале происходит связывание фактора транскрипции со специфической последовательностью ДНК. Этот фактор находит свое место на хроматине. Далее происходит присоединение комплекса ремоделирования хроматина и изменяется структура нуклеосом. Связывание комплекса ацетилирования и ковалентная модификация гистонов свидетельствуют о том, что произошла активация локуса.
В области промотора также происходит модификация ДНК. Метилирование цитозина, входящего в состав дублетов CpG, связано с инактивацией гена. Пока не очень понятно, почему ДНК становится мишенью метилирования.
Очевидно, что для ремоделирования хроматина в области промотора необходимо прохождение различных событий, включающих ацетилирование, в результате которых нуклеосомы изменяются. Однако какие изменения необходимы в гене, чтобы обеспечить прохождение РНК-полимеразы? Известно, что РНК-полимераза может транскрибировать ДНК in vitro со скоростью, близкой к скорости in vivo (25 нуклеотидов в сек), но только на матрице свободной ДНК. Охарактеризованы белки, способные увеличивать скорость, с которой РНК-полимераза транскрибирует хроматин in vivo. Все они действуют на хроматин.
Современное представление о механизме их действия заключается в том, что они связываются с РНК-полимеразой и движутся вместе с ней вдоль матрицы, изменяя структуру нуклеосом за счет модификации гистонов. К числу этих белков относится ацетилаза гистонов. Возможно также, что первая молекула РНК-полимеразы, транскрибирующая ген, играет роль «пионера» и содержит факторы, изменяющие структуру единицы транскрипции таким образом, чтобы сделать ее более доступной для последующих молекул фермента.
Комплексы, модифицирующие структуру или активность хроматина, характеризуются наличием субъединиц-мишеней,
которые определяют точки связывания с ДНК, т. е. мишени для HAT или HDAC ферментов,
ацетилирующих или деацетилирующих гистоны, и эффекторных субъединиц, обладающих другим действием на хроматин или ДНК.
Активация промотора включает этапы связывывания активатора,
специфичного по отношению к определенной последовательности,
отбор и функционирование ремоделируемого комплекса, а также комплекса ацетилирования