Диацилглицерол- и инозитол-1,4,5-трифосфат-связанные рецепторы. ДНК-связанные рецепторы
Многие G-белки активируют диацилглицерол- и инозитол-1,4,5-трифосфатный путь. Один из G-белков, обозначаемый Gq, стимулирует активность фосфолипазы С. Этот фермент, в свою очередь, способствует продукции диацилглицерола (ДАГ) и инозитол-1,4,5-трифосфата (IР3) через гидролиз полифосфотидинозитов. Альтернативный путь включает активацию G-белками мембранной фосфолипазы А2, что ведет к образованию ДАГ и фосфатидной кислоты. Эти компоненты трансдукции обладают различными эффектами.
IР3 представлен не только инозитолфосфатом, продуцируемым фосфолипазой в клетках. Существует множество соединений, которые могут обладать различными функциями. Считается, что инозитол-1,3,4,5-тетрафосфат облегчает вход кальция в различные внутриклеточные компартменты.
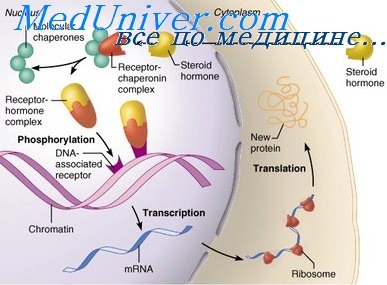
ДНК-связанные рецепторы
Внутриклеточные рецепторы, которые взаимодействуют с ДНК, связываются с гормонами, такими как ретиноевая кислота, кортикостероиды, тиреоидине гормоны и витамин D. Эти рецепторы состоят из ядерных белков. Таким образом, агонисты должны пройти через клеточную мембрану, чтобы достичь рецептора. Например, стероиды проникают в клетку и связываются с цитоплазматическим рецептором, который зачастую имеет прицепленную тормозящую молекулу, например белок теплового шока с массой 90 кДа (HSP90). Молекулярным ответом является изменение конформации рецептора, что ведет к диссоциации рецептора от тормозящей молекулы. Существует множество вариантов клеточных ответов на активацию ДНК-рецепторов.
Действие агонистов на тирозинкиназные рецепторы сопряжено с регуляцией роста, клеточной дифференцировкой и ответом на метаболические стимулы. К эндогенным агонистам относят инсулин, фактор роста эпидермиса и фактор роста тромбоцитов. Агонисты вызывают изменение конформации рецептора и действуют как тирозинкиназные ферменты, фосфорилируя остатки тирозина в широком спектре молекул внутри клетки.
Рецептор-связанные каналы
РСК состоят из субъединиц, каждая из которых имеет четыре трансмембранных домена. Эти домены образуют комплексы с различной стехиометрией. Существует несколько различных типов РСК. Каждый несет в своей структуре внеклеточный центр, связывающий лекарство. При присоединении лекарства к нему запускаются конформационные изменения других частей молекулы РСК, что приводит к открытию центральной ион-селективной поры. Ток ионов происходит только при открытой поре. Молекулярная конфигурация канала в этом случае определяет состояние канала.
Простейшая модель состояний канала включает три варианта:
• состояние покоя (неуправляемый канал, т.е. закрытый, но открывающийся в ответ на соответствующий стимул);
• состояние активации (открытый канал);
• состояние инактивации (т.е. закрытый и неспособный к открытию канал в ответ на соответствующий стимул в отличие от канала в состоянии покоя).
Некоторые лекарства обладают молекулярным механизмом действия, который вовлекает модуляции перехода РСК из одного состояния в другое.
Центр, связывающий лекарство, обычно обозначают как рецептор (который оперирует каналом). В данной книге РСК обозначен значком, отличающимся от значка для рецепторов гормонов, нейротрансмиттеров и аутакоидов. РСК для удобства обозначают как рецептор, а также канал или канал с рецепторным центром. Так, никотиновый холинергический рецептор является РСК (этот РСК содержит два центра, связывающих лекарство).
В большинстве случаев агонист РСК открывает канал, в то время как антагонист препятствует его открытию, а обратный агонист закрывает открытый канал. К РСК относят:
• никотиновые РСК (активируются ацетилхолином);
• ГАМК-РСК (активируются гамма-аминомасляной кислотой, ГАМК);
• глициновые РСК (активируются глицином);
• 5-НТ3-РСК (активируются 5-гидрокситриптамином);
• Р2х-РСК (активируются аденозином).