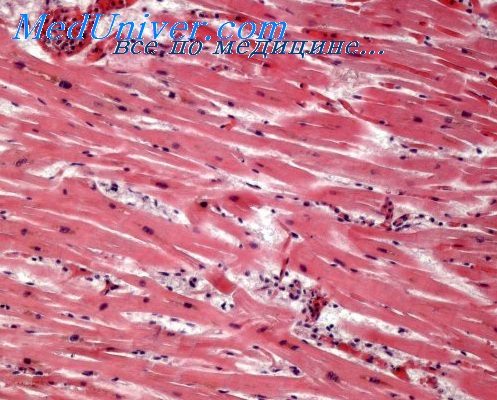
Миофиброз при врожденных пороках. Компенсаторная гипертрофия миокарда
Миофиброз в правом желудочке при тетраде Фалло и в левом желудочке при врожденном сужении устья аорты в ряде случаев является диффузным, поскольку подвергшиеся коллагенизации решетчатые волокна пронизывают всю толщу стенки наиболее гипертрофированного желудочка. Тем не менее самой большой степени процесс коллагенизации решетчатых волокон достигает во внутренних слоях миокарда, в трабекулярных и папиллярных мышцах.
Таким образом, аргирофильная строма гипертрофированного сердца при врожденных пороках изменяется по тем же законам, что и при гипертрофии сердца другой этиологии.
Паренхима и строма сердца представляют собой неразрывное целое. Понятие «компенсаторная гипертрофия миокарда» включает в себя и понятие «гиперплазия стромы», так как гипертрофия миокарда при различных видах врожденных пороков, а также при гипертрофии сердца другой этиологии сопровождается разрастанием сети продольных и поперечных интрамуральных решетчатых волокон.
О значении соединительнотканной основы мышечной ткани свидетельствует работа В. А. Энгельгардта (1948), в которой автор указывает, что движущей силой сокращения служит эластическое натяжение растянутой соединительнотканной основы, поэтому активной фазой мышечной деятельности является фаза расслабления. Миозиновая структура в мышечном волокне обусловливает сопротивление тянущей силе соединительнотканной основы.
Автор считает, что эластичность сокращенного и растянутого мышечного волокна зависит исключительно от оплетающей это волокно соединительнотканной стромы.
Мы не будем подробно разбирать очень важные и интересные вопросы фибриллогенеза в гипертрофированном миокарде, поскольку они специально обсуждены нами ранее (1949).
Следует отметить, что вопросы фибриллогенеза все еще далеки от решения, поскольку даже электронная микроскопия не дала этой возможности (В. П. Гилев, 1959). Укажем лишь, что нами особенно замечены высказывания некоторых гистологов о том, что волокна соединительной ткани могут быть образованы не только соединительнотканными клетками, но и всеми структурами мезодермального происхождения, в том числе гладкомышечными и поперечнополосатыми волокнами [Даблин (Dablin, 1946)]. М. Ф. Глазунов (1933, 1934) доказал образование аргирофильных волокон в виде тончайшей войлокообразной сети на поверхности протоплазмы клеток в опухоли из поперечнополосатой мускулатуры и в новообразовании из гладкомышечных элементов.
Мы, так же как М. Ф. Глазунов (1947), допускаем, что мышечные волокна обладают фибриллогенетической способностью, что связано с генетическим родством соединительнотканных и мышечных волокон в смысле их единой мезенхимальной природы.
Даже сторонники эктоплазматической теории образования волокнистых структур соединительной ткани принимают положение о фибриллогенетической способности мышечных элементов (Г. В. Ясвоин, 1935).
А. А. Заварзин (1947) указывает, что «образование опорных структур является элементарным свойством животной клетки, и притом не только соединительнотканной, но и эпителиальной». Н. Г. Хлопин (1946) считает «способность к образованию межклеточных веществ — одним из элементарных свойств живой протоплазмы, быть может, одной из форм ее взаимодействия с окружающей средой». Он указывает, что «аргирофильные волокна могут быть образованы клетками различного происхождения, а не только производными мезенхимы».
Особенно интересно то, что в эксплантате из мускулатуры сердца отмечается новообразование решетчатых волокон (В. Цымбал, 1938).