Hodgkin А о возбуждении кардиомиоцитов. Фазы возбуждения мышечных клеток
Hodgkin А. (1952 — 64) показал, что при возбуждении происходит не простая нейтрализация мембраны, как предполагал Bernstelng, а ее перезарядка (реверсия) с накоплением положительных ионов внутри клетки и освободившихся отрицательных ионов снаружи. При этом потенциал внутренней поверхности мембраны достигает +20 +30 мВ. Однако, такой ТП держится очень короткое время, т.к. прекращается ток Na+ по быстрым каналам в клетку и возникает (фаза 1) ток отрицательных ионов С1, что резко уменьшает внутриклеточный потенциал до 0. Медленные токи Na+ и Са++ в клетку и К+ из клетки обуславливают фазу 2 (плато) ТП действия (рис. 3), знаменующую поэтапное окончание деполяризации и начало реполяризации — фазу медленной реполяризации. В фазу ЗТП происходит быстрая реполяризация и полное восстановление исходной поляризации мембраны к фазе 4 (ТП покоя — ДТП).
Для восстановления исходного химического состава в клетке и окружающей ее жидкости все время возбуждения в период систолы и после его угасания (диастола) действует «натриевый и кальциевый насос», выводящий Na+ и Са++ из клетки, т. е. против их концентрационных градиентов. Это возможно благодаря энергии метаболических процессов, главным образом синтеза АТФ. Одновременно К+ входит в клетку в количестве приблизительно равном выходящему Na+. Постепенно восстанавливается исходный химический состав клетки и она готова к следующему циклу возбуждения и сокращения.
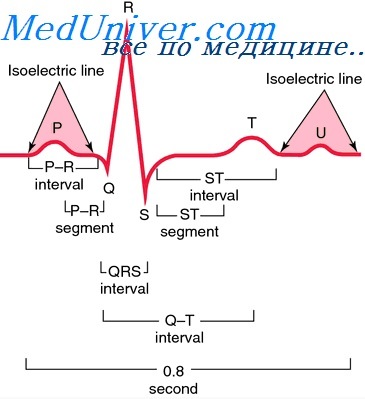
Описанный механизм возбуждения клетки свойственен большинству возбудимых тканей, в том числе сократительным волокнам сердечной мышцы. Однако характерной особенностью возбуждения последних по сравнению с другими клетками является кратковременность периода «быстрой» деполяризации и большая продолжительность периода реполяризации. Общая продолжительность ТП действия сократительного кардиомиоцита желудочков меньше чем в специализированных проводниковых волокнах атриовент-рикулярной системы Гиса-Пуркинье. Реполяризация последних запаздывает по отношению к реполяризации сократительных кардиомиоцитов (фазе 3) и обуславливает наличие небольшого потенциала в начале фазы 4 соответствующего на ЭКГ зубцу U (Гоффман Б. и Крейнфилд П., 1962).
Закономерность распространения возбуждения по нервному и мышечному волокнам была обоснована в теории «локальных токов» Ходжкина. Согласно этой теории, возбуждение распространяется от возбужденного участка волокна к покоящемуся вследствие прохождения тока, генерируемого при возбуждении участка. Этот ток понижает потенциал клеточной мембраны непосредственно перед активируемым участком, уменьшая накопленный заряд. Снижение мембранного потенциала ведет к повышению проницаемости мембраны для натрия. Последний создает положительный заряд внутренней поверхности мембраны и отрицательный — наружной поверхности на новом участке. При этом генерируется электрический ток. понижающий потенциал следующего участка мембраны. Так волна перезарядки распространяется по волокну, вовлекая в возбуждение все новые и новые участки.
Динамика трансмембранного потенциала автоматических Р-клеток существенно отличается от ТП сократительных клеток как в период электрической систолы, так и, особенно, в период диастолы. Автоматизм, т. е. воспроизводство импульсов к возбуждению, связано прежде всего с наличием в ТП Р-клеток спонтанной диастолической деполяризации (СДД) — изменения величины ТП в течение диастолы.
Т. е. в течение фазы 4 отрицательный трансмембранный потенциал Р-клеток снижается с -70 мв до порогового потенциала (ПП) равного -55 мв, после чего возникает импульс — относительно медленная фаза 0 ТП действия. Автоматические клетки (центр автоматизма) обладающие наибольшей скоростью диастолической деполяризации (т е. раньше других Р-клеток достигающие порогового потенциала) становятся водителем сердечного ритма — номотопным центром автоматизма, т. к. их импульс распространяясь по миокарду разряжает потенциальные (зреющие) импульсы других (гетеротопных, эктопических) центров автоматизма т. е. прерывает спонтанную диастолическую деполяризацию в Р-клетках гетеротопных центров.
Видео физиология фаз потенциала действия кардиомиоцитов