Большая часть (до 80-90%) внутриклеточного распада белков осуществляется убиквитин-протеасомным путем. Убиквитин-протеасомный путь присутствует в ядре и цитоплазме эукариотических клеток и играет важную роль в распаде нормальных и аномальных белков. Этот путь отвечает за регулируемое расщепление многих белков, в том числе тех, которые необходимы для контроля роста и пролиферации клеток, клеточной дифференцировки, иммунных и воспалительных реакций, апоптоза и метаболической адаптации.
Убиквитин-протеасомный путь также осуществляет «хозяйственные» функции в основном кругообороте белка и элиминации аномальных белков с неверной кодировкой, неправильно свернутых, локализованных в несвойственных им местах, поврежденных или иным образом выведенных из действия. Убиквитин-протеасомный путь играет решающую роль в контроле мышечной массы, и его активность повышена при кахексии. Этот путь также играет существенную роль в восстановлении мышц и их ремоделировании.
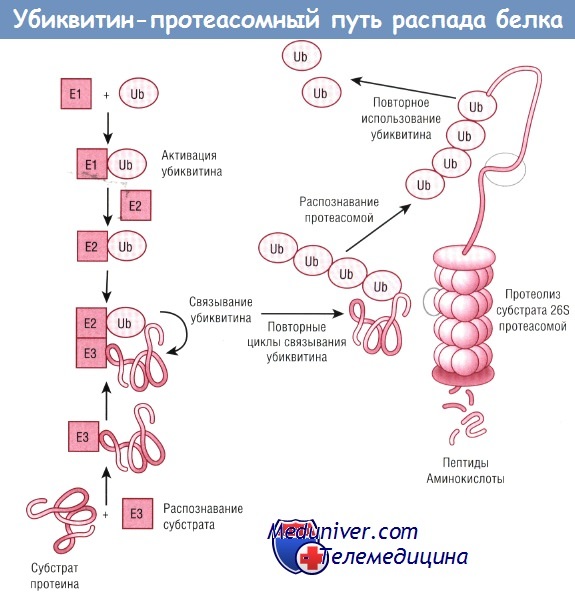
Убиквитин-протеасомный путь можно рассматривать как последовательность трех процессов:
(1) распознавания белкового субстрата для распада;
(2) ковалентного присоединения цепочки полиубиквитина в качестве метки белка для распада;
(3) протеолиза белка комплексом 2500 кДа, называемым протеасомой 26S.
Распознавание белка, предназначенного для распада, обычно осуществляется:
(1) по определенным структурным изменениям белка, в том числе по воздействию на определенную последовательность аминокислот, которая в норме скрыта посттрансляционными модификациями, такими как фосфорилирование или гидроксилирование;
(2) по присоединению или высвобождению его лигандов;
(3) по взаимодействию с белком-адаптером или шапероном (например, экспорт неправильно свернутых белков с помощью шаперонов из ЭПР в цитозоль);
(4) по специфическому повреждению, произошедшему в белке в результате окисления или нитрозилирования.
Кроме того, наличие специфических «дестабилизирующих» остатков на iV-концевом участке маркирует пептид, предназначенный для расщепления (т.е. с более коротким периодом полураспада). Однако следует отметить, что перед тем, как подвергнуться расщеплению, не все белки получают убиквитиновую метку.
Напротив, некоторые белки проходят расщепление с помощью 20S основных протеасом. Цель этой модели расщепления неясна, однако это, по-видимому, происходит с белками, имеющими отчетливо неструктурированные участки, придающие белку большую нестабильность.
Распад белка убиквитин-протеасомным путем является тщательно регулируемым многоступенчатым процессом.
Первая реакция — активация убиквитина Е1 (убиквитинактивирующим ферментом), после чего убиквитин доставляется к Е2 (убиквитинприсоединя-ющему ферменту).
Вторая реакция — образование комплекса с Е2, ЕЗ (убиквитин-связывающим ферментом) и субстратом.
Третья реакция — передача убиквитинов лизину(-ам) субстрата, чтобы пометить субстрат полиубиквитиновой цепочкой. В результате четвертого шага полиубиквитиновый субстрат отделяется от Е3.
Наконец, протеасома разворачивает субстрат АТФ-зависимым способом, за счет активности убиквитингидралазы отделяет убиквитиновую цепочку и вставляет развернутый белок в протеасомную камеру, где находятся участки протеазной активности. Молекулы убиквитина используются вторично, в то время как образовавшиеся пептиды расщепляются до аминокислот и используются повторно для синтеза нового белка.
Ub — убиквитин.
После того как белок был определен в качестве субстрата для деградации, он ковалентно снабжается меткой убиквитина. Убиквитин — белок, присутствующий во всех типах клеток и состоящий из 76 аминокислотных остатков, в том числе С-концевого глицина и лизинового остатка в положении 48. Убиквитин ковалентно связан с белком, предназначенным для деградации в серии из трех реакций, катализируемых ферментами, известными как Е1 (убиквитинактивирующий фермент), Е2 (убиквитинприсоединяющий фермент) и Е3 (убиквитинсвязывающий фермент).
Существуют две изоформы Е1, несколько изоформ Е2 и очень большое количество ферментов Е3, что позволяет осуществлять специфическое для многих тканей и субстратов регулирование этого процесса.
Сначала молекула убиквитина активируется путем связывания с Е1 в АТФ-зависимой реакции, затем фрагмент убиквитина передается к Е2. И Е1, и Е2 имеют остатки активных участков цистеинила, которые образуют тиоэфиры с С-концевым глициновым остатком убиквитина. Наконец, убиквитин, присоединенный к Е2, передается напрямую или через Е3 к внутреннему лизиловому остатку на субстрате белка.
Е3 играет важную роль в распознавании субстрата белка для расщепления и в регулировании образования субстратного комплекса Е2/Е3. Аналогичным способом дополнительные молекулы убиквитина присоединяются к субстрату, имеющему одну присоединенную молекулу убиквитина, образуя изопептидные связи между С-концевым глициновым остатком молекулы убиквитина и лизином, расположенным в положении 48 ранее добавленной молекулы убиквитина.
Необходима цепочка по крайней мере из четырех молекул убиквитина для того, чтобы белки, помеченные несколькими молекулами убиквитина, были с легкостью распознаны и направлены к протеасоме 26S для расщепления. Недавно был выполнен обзор связывающих доменов и активности убиквитина. Важно отметить, что процесс убиквитинации является обратимым, с наличием процесса отсоединения, который совершается классом цистеиновых протеаз, называемых деубиквитинирующими ферментами.
Деятельность деубиквитинирующего фермента заключается в устранении связывания с убиквитином, обработке предшественников убиквитина, компоновке убиквитиновых цепей и повторном использовании убиквитина. Эти процессы отвечают за регулирование нескольких сигнальных путей, важных для процесса развития, включая рост и дифференцировку клеток.
Фактическое расщепление убиквитинированных белков происходит во внутренней камере протеасомы, но молекулы убиквитина отщепляются первыми, поэтому могут быть использованы повторно, протеасома 26S является большим, состоящим из нескольких субъединиц комплексом, в который входит субъединица 20S в качестве протеолитического ядра с регулирующим комплексом 19S, присоединенным к одному или обоим концам. Регулирующие субъединицы участвуют в распознавании меченых белков, удалении убиквитиновых меток, а также в АТФ-зависимых процессах разворачивания белка и направления его в протеолитическое ядро, имеющее форму туннеля.
Протеолитический комплекс 20S представляет собой похожую на бочонок структуру, состоящую из четырех уложенных одна на другую кольцевых структур (abba), каждая из которых образована семью субъединицами. Центральная каталитическая полость данной структуры содержит в общей сложности шесть протеолитических участков, образованных тремя отдельными каталитическими субъединицами каждого b-кольца. Эти каталитические субъединицы относятся к TV-концевым треониновым гидралазам, поскольку iV-концевой треонин выступает в качестве нуклеофильного катализатора. Однако три различные субъединицы в каждом из двух колец отличаются предпочтениями в расщеплении пептидных связей сразу после основных, гидрофобных или кислых остатков.
Субъединица 20S гидролизует входящий субстрат на пептидные фрагменты, состоящие из 3-30 аминокислотных остатков. Эти пептидные продукты высвобождаются из протеасомы и далее в клетке подвергаются гидролизу другими протеазами и аминопептидазами.
Регулирование протеолиза в протеасоме происходит на трех уровнях. Во-первых, распознавание субстрата регулируется функциями, которые устанавливают целевой белок для проведения полиубиквитинирования. Для большинства белков субстраты являются неопознанными и включают фосфорилирование, гидроксилирование остатка пролина или выявление сигнала для расщепления, содержащегося в первичной последовательности. Во-вторых, регулируемая деградация конкретных классов субстрата может быть достигнута путем объединения комплексов Е2/ЕЗ с различными вспомогательными факторами.
Например, в некоторых случаях именно ЕЗ должен быть изменен или «включен» посредством посттрансляционной модификации, чтобы принять активную форму, которая распознает субстрат. В других случаях стабильность белкового субстрата зависит от его связи с молекулярными шаперонами, которые действуют как распознающие элементы и служат в качестве соединения с соответствующими лигазами. В частности, инсулин уменьшает убиквитин-опосредованную активность протеасомы путем перемещения внутриклеточных протеаз и инсулиноснижающего фермента от протеасом 20S и 26S. Наконец, убиквитин-протеасомный путь можно регулировать посредством взаимодействия либо изменения экспрессии убиквитина или протеасомных субъединиц.
Пример этого можно наблюдать после внутривенной инфузии аминокислот или после введения их в просвет кишечника. Увеличение поставки аминокислот, но не глюкозы снижает экспрессию мРНК убиквитина, убиквитинприсоединяющего фермента с молекулярной массой 14 кДа, и С9-субъединицы протеасомы в слизистой оболочке кишечника.