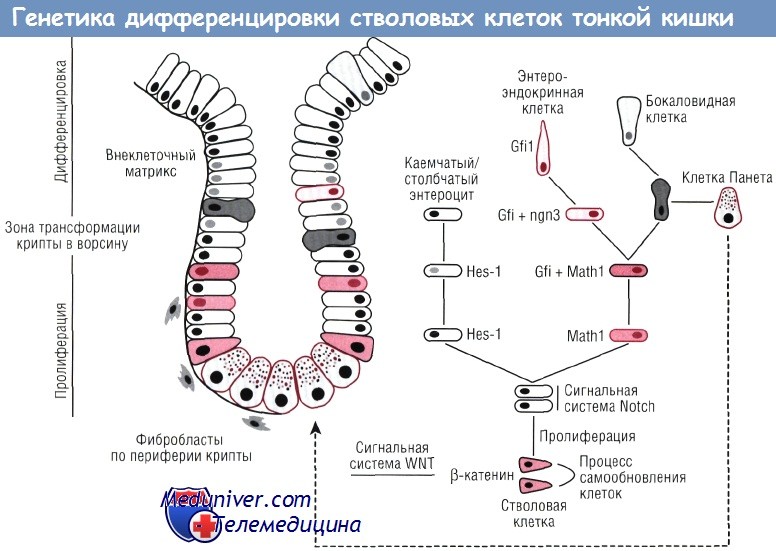
Значение сигнального пути WNT в регуляции стволовых клеток тонкой кишки
Рассмотрение вопроса об основах молекулярной регуляции стволовых клеток и последующего распределения клеточных популяций следует начать с процесса нокаута гена Tcf-4 как важного медиатора сигнального пути WNT в кишечнике. Белки системы WNT представляют собой секреторные гликопротеины, участвующие в дифференцировке и органогенезе многочисленных тканей. У мышей с нокаутом гена Tcf-4 отмечается исчезновение пролиферирующих эпителиальных клеток на поздних стадиях внутриутробного развития.
Следовательно, ген Tcf-4 представляет собой основной регулятор функции стволовых клеток. При связывании клеток с белками WNT из внутриклеточного комплекса высвобождается b-катенин вместе с белком, ассоциированным с аденоматозным полипозом кишечника (АРС), что активирует ген Tcf-4 и приводит к пролиферации клеток. При отсутствии воздействия со стороны WNT ген Tcf-4 выступает как ингибитор пролиферации, и происходит дифференцировка клеток. Две группы исследователей изучали угнетение сигнального пути WNT с использованием ингибитора dickkopf (DKK), блокирующего поверхностные клеточные рецепторы, либо на трансгенных мышах, либо с использованием вирусного вектора.
Если сигнальный путь WNT блокировали действием DKK, то пролиферация прекращалась и крипты исчезали. При элиминации ингибитора происходили восстановление пролиферативных процессов и регенерация крипт. Представленные данные свидетельствуют о том, что сигнальный путь WNT регулирует процессы клеточной пролиферации, однако он не является необходимым для поддержания жизнеспособности стволовой клетки в течение периода ее формирования. Напротив, элиминация у лабораторных мышей белка АРС, который в нормальных условиях способствует связыванию b-катенина в цитоплазме, приводила к накапливанию данного белка в клеточном ядре и активации сигнального пути WNT во всем эпителии.
Все эпителиальные клетки, расположенные как в криптах, так и на поверхности ворсин, трансформировались в пролиферирующие недифференцированные клетки. Более того, при утрате белка АРС в эпителии также нарушалась экспрессия мезенхимой компонентов внеклеточного матрикса. В недавно опубликованном исследовании, где инактивация белка АРС достигалась несколько иным способом, также обнаружено, что зона пролиферации чрезмерно расширялась за счет пролиферации дифференцированных клеток кишечных ворсин. Кроме того, увеличивалось количество клеток Панета вследствие повышенной экспрессии генов, управляемых сигнальным путем WNT. Аналогичная функция сигнального пути WNT отмечена группой исследователей под руководством van Es.
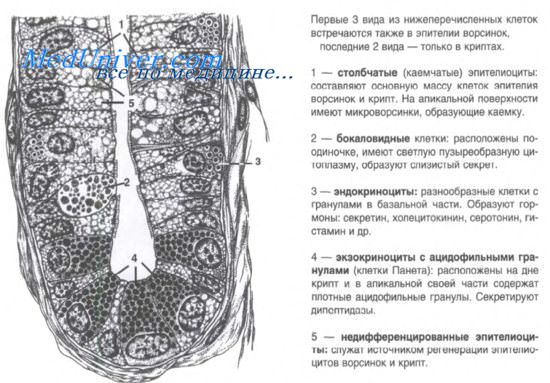
Клетки крипты тонкой кишки
Указанные исследования свидетельствуют, что сигнальный путь WNT раздельно управляет программой пролиферации стволовых клеток и программой созревания клеток Панета. Механизм данного процесса, однако, точно не установлен. В то же время имеющиеся научные факты позволяют предположить, что бокаловидные клетки и клетки Панета происходят из одной клеточной линии.
Непосредственно эпителий крипт секретирует белки WNT-6 и WNT-9b. Всесторонний анализ характера экспрессии матричной РНК (мРНК) позволил установить наличие в эпителии крипт рецепторов frizzled-5 и frizzled-6 при одновременном синтезе в окружающей мезенхиме антагониста рецептора — sFRP-1. Клетки в базальной пластине ворсин экспрессируют мРНК, необходимую для синтеза WNT-2b, WNT-4 и WNT-5a. Clevers и соавт. предложили модель, в соответствии с которой эпителий крипт имеет определенный градиент WNT-факторов. Их высокое содержание, как известно, стимулирует клеточную пролиферацию. По мере миграции клеток в направлении от основания крипты уровень WNT-факторов снижается, что приводит к торможению процессов клеточной пролиферации и стимулирует процессы дифференцировки.
Полученные в последних исследованиях результаты сопоставимы с принципами данной модели, где градиенты белков WNT, рецепторов к компонентам сигнального пути WNT, и WNT-антагонисты управляют стволовыми клетками тонкой кишки и процессами пролиферации и дифференцировки эпителиальных клеток.
Кроме того, многочисленные клеточные белки, роль которых до настоящего времени не выяснена, вовлечены в работу сигнального пути WNT. EphB2 и EphВЗ под воздействием белка Tcf-4 проявляют ингибирующие свойства и, как было продемонстрировано, являются крайне важными компонентами в регуляции процесса миграции эпителиальных клеток. Белки Eph и ephrin представляют собой трансмембранные сигнальные белки, необходимые для процессов межклеточной адгезии и сортировки клеток. У мышей при отсутствии продукции как EphB2, так и EphВЗ клетки Панета мигрируют к верхушке ворсины в большей степени, чем в направлении основания крипты.
Соответственно, нокаут b-катенина, регулирующего белок EphВЗ, приводит к тому, что клетки Панета располагаются аномально. Эти данные свидетельствуют о том, что созревание клеток Панета представляет собой автономный клеточный процесс, а не результат распределения и сортировки клеток в кишечной крипте, которые контролируют сигнальный путь Eph/ephrin. Вероятно, нарушение сигнальных механизмов не сопровождается нарушением процесса клеточной дифференцировки и формирования кишечных ворсин в целом во время внутриутробного развития. Предполагается, что данный процесс контролируют другие механизмы.