Регуляция специализации энтодермы при формировании желудочно-кишечного тракта у плода
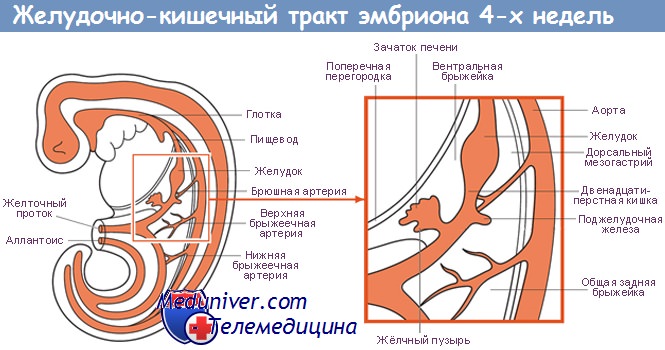
Развитие ЖКТ происходит в три этапа. Первый этап — первичная специализация энтодермы. Второй этап заключается в формировании и структурировании кишечной трубки, что определяет передне-заднюю ось эмбриона и границы между различными органами. На третьем этапе начинается образование органов, которые в процессе развития выходят за границы кишечной трубки, например печени и поджелудочной железы.
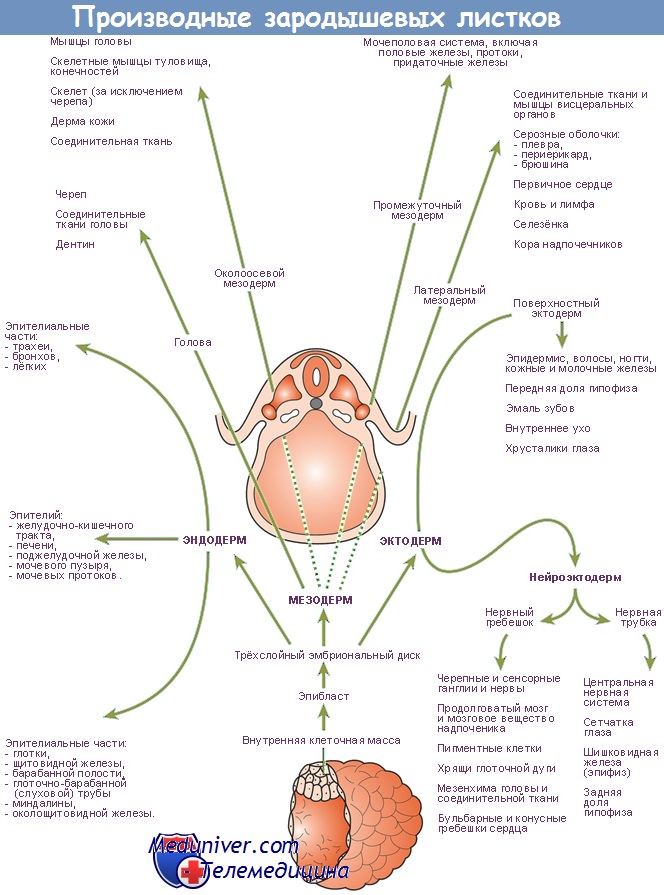
В экспериментах на лабораторных животных установлены эволюционно консервативные группы генов, принимающих участие в специализации энтодермы, и ряд других генов, участвующих только в процессе развития ЖКТ позвоночных. Эпителий этого тракта формируется из энтодермы — одного из эмбриональных зародышевых листков, возникающих в процессе гаструляции. Исследования, проведенные на таких лабораторных организмах, как круглые черви (почвенные нематоды Caenorhabdilis elegans), плодовые мушки (род Drosophila), африканские шпорцевые лягушки (род Xenopus), аквариумные рыбки Danio rerio и лабораторные мыши, позволили установить некоторые основные молекулярные механизмы регуляции процесса развития энтодермы. Моделирование на лабораторных мышах, у которых был проведен нокаут определенных генов для исследования их функций, в настоящее время рассматривается в качестве основы для изучения таких заболеваний ЖКТ у человека, как стеноз привратника, различные атрезии, неперфорированный анус.
Специализация энтодермы может быть прослежена на самых ранних стадиях развития эмбриона. В классических экспериментальных исследованиях продемонстрирована способность эксплантатов куриных эмбрионов еще до процесса гаструляции, в культуре, к развитию ЖКТ, что свидетельствует о четкой детерминации процесса уже на данном этапе эмбрионального развития. Энтодерма куриных эмбрионов, выделенная и помещенная в лабораторных условиях в культуру до формирования ЖКТ, характеризуется экспрессией молекулярных маркеров, которые должны в норме появиться in vivo, что свидетельствует о наличии феномена ранней специализации.
Появляются доказательства в поддержку гипотезы, что отличительные особенности энтодермы обусловлены функциональной клеточной независимостью, однако для нормального развития органов необходимо тесное взаимодействие энтодермы и мезодермы. В настоящее время на многочисленных лабораторных организмах установлены целые группы генов, способствующих специализации энтодермы. Один класс генов кодирует факторы транскрипции, которые, в свою очередь, непосредственно активируют гены-мишени. Второй класс генов кодирует синтез сигнальных молекул, обеспечивающих процесс межклеточной кооперации. Как минимум несколько факторов транскрипции, участвующих в процессе специализации энтодермы, продолжают экспрессироваться в тканях ЖКТ в течение всего последующего его развития, например семейства генов Fox и факторов GATA (семейство факторов транскрипции, характеризующихся способностью связываться с фрагментом дезоксирибонуклеиновой кислоты (ДНК) и с GATA-последовательностью нуклеотидов).
В регуляции развития ЖКТ принимают участие, действуя в строго определенное время в различных локусах, сигнальные пути, или механизмы передачи регуляторных стимулов внутри клетки через определенную цепь молекул-посредников, представители группы факторов роста — трансформирующие факторы роста b в том числе непосредственно трансформирующий фактор роста b (TGF-b), и белок морфогенеза костной ткани (BMP), а также сигнальные пути hedgehog (сигнальные системы млекопитающих, представленные тремя различными белками). Продемонстрировано, что фактор транскрипции Sox-17 играет крайне важную роль в развитии энтодермы у мышей на ранних этапах эмбриогенеза, однако роль Sox-17 в дальнейшем не установлена. В последнее время выявлено, что сигнальный путь Notch (рецепторзависимый латентный фактор транскрипции, локализованный в клеточной цитоплазме) также выполняет важную функцию в регуляции развития ЖКТ.
До настоящего времени не удалось выяснить, существует ли «главный ген», запускающий формирование энтодермы и последующее развитие ЖКТ. При исследовании некоторых животных моделей установлены гены, присутствие которых является необходимым и в то же время достаточным условием для формирования энтодермы, например ген mixer у шпорцевых лягушек. Ген мышей mixl1 играет основную роль в дифференцировке эмбриональных клеток в энтодерму. У других лабораторных животных обнаружены гены, необходимые для обеспечения данного процесса, однако недостаточные для формирования полноценной энтодермы.
В исследованиях на лабораторных животных установлены начальные этапы индукции образования энтодермы. Энтодерма у мышей формируется из передней части первичной полоски эпибласта. Ключевой регулятор ранних этапов развития энтодермы — nodal-лиганд факторов роста суперсемейства TGF-b. Nodal экспрессируется в расположенном в бластодерме организационном центре или в узелке первичной бороздки эмбриона мыши. В отсутствие экспрессии nodal эмбрионы мыши не способны к гаструляции или к формированию дифинитивной энтодермы. Аналогично у эмбрионов Danio rerio в отсутствие экспрессии гомологов nodal также не образуется энтодерма. Напротив, экспрессия функционально активных рецепторов к TGF-b способствует преобразованию эмбриональных клеток в энтодерму. Реализация сигнального пути TGF осуществляется посредством стабильного комплекса промежуточных белков Smad.
Необходимым для развития энтодермы является белок Smad2, и нокаут этого белка приводит к нарушению формирования энтодермы задней кишки, а разные мутации в гене, кодирующем Smad2, вызывают различные аномалии передней/головной кишки. FoxA2 также играет важную роль в развитии энтодермы, являясь мишенью сигнального пути, запущенного фактором nodal через белок Smad2. Smad2 и Smad3 совместно регулируют процесс развития энтодермы у мышей, оказывая различное влияние на формирование передней кишки и задней кишки. Кроме того, сигнальная система Smad необходима для развития печени. Основную роль в формировании печени играет ген Hex, экспрессия которого отсутствует либо существенно снижена при нарушенной продукции белков Smad.
Этапы развития желудочно-кишечного тракта у плода
Этап
Минимальный срок гестации
Гаструляция
3-я неделя
Формирование первичной кишки; закладка печени и поджелудочной железы
4-я неделя
Рост кишечной трубки
7-я неделя
Образование кишечных ворсин
8-я неделя
Погружение первичной кишки в брюшную полость
10-я неделя
Завершение органогенеза
12-я неделя
Появление обкладочных клеток желудка, формирование островков поджелудочной железы, секреция желчи, появление кишечных ферментов
12-я неделя
Появление глотательных движений
16—17-я недели
Функциональная зрелость
36-я неделя
Установлено, что энтодерма находится в тесном контакте с мезодермой на протяжении всего ЖКТ. В экспериментальных исследованиях, выполненных на культурах тканей, было показано, что формирование энтодермы и ее дифференцировка в отдельные органы представляет собой результат взаимодействия мезодермы и энтодермы. Раньше всего было показано, что для формирования переднезадней оси энтодермы мышей необходимо сигнальное воздействие мезодермы на энтодерму посредством фактора роста фибробластов (FGF).
Другие члены семейства FGF и рецепторы к ним необходимы для формирования печени. Еще три семейства генов крайне необходимы в процессе взаимодействия мезодермы и энтодермы — гены, кодирующие факторы сигнальных путей hedgehog и BMP, гены box.
У почвенной нематоды Caenorhabditis elegans для специализации клеток, из которых развивается кишечный эпителий, необходимы два гена, кодирующие факторы транскрипции GATA, в то время как фактор GATA у плодовых мушек рода Drosophila кодируется одним геном serpent, наличие которого нужно для дифференцировки кишечной энтодермы. В кишечнике позвоночных экспрессируются три фактора группы GATA. Предполагается, что в процессах пролиферации и дифференцировки кишечного эпителия факторы GATA4, -5 и -6 выполняют различные функции, однако их функциональное значение на ранних этапах развития кишечной трубки у млекопитающих пока не установлено. Помимо факторов GATA, представители семейства forkhead-related факторов (кодируемые генами Fox) и сигнального пути WNT/Tcf являются важными регуляторами развития энтодермы.
У позвоночных важную роль в инициации формирования энтодермы играют представители суперсемейства TGF-b. Молекула цитоскелета ELF3 задействована в сигнальном пути через факторы TGF-P, а также необходима для развития энтодермы. В отсутствие гена, кодирующего ELF3, у мышей полностью отсутствует кишечная энтодерма.
Многочисленные факторы транскрипции, первоначально считавшиеся специфическими для формирования печени, играют роль и в развитии кишечника. При изучении эмбрионов мыши было отмечено, что некоторые из факторов транскрипции имеют характерные паттерны экспрессии, что позволило предположить их участие в развитии кишечника. Например, нуклеарный печеночный фактор (HNF) 3b (сейчас известный как FoxA2) оказался важным элементом в процессе самых ранних этапов дифференцировки ЖКТ и продолжает экспрессироваться в производных энтодермы у взрослых. Гомозиготные мутанты с полным отсутствием HNF3b не способны к формированию первичной полоски, предшествующей образованию кишечной трубки и ряда других структур. HNF3b участвует в образовании передней и средней, но не задней кишки.
Идентифицировано большое количество факторов семейства Fox, некоторые из которых характеризуются преимущественной или специфической экспрессией в кишечнике. Один из представителей данного семейства, Foxl1, в норме экспрессируется кишечной мезодермой и представляет собой основной медиатор взаимодействия между эпителием и мезенхимой. Отсутствие указанного фактора сопровождается патологической пролиферацией эпителиальных клеток и нарушением развития кишечника. Этот фактор действует через экспрессию протеогликанов, проявляющих себя как корецепторы к компонентам сигнального пути WNT, и таким образом активирует комплекс WNT/P-катенин, регулирующий пролиферацию клеток. Группа исследователей во главе с Zaret предложила модель, в которой факторы FoxA2 и GATA открывают ДНК в клетках энтодермы, обеспечивая ее доступность для последующего связывания с факторами транскрипции, регулирующими клеточно-специфические гены. Таким образом, вероятно, при развитии кишечника многочисленные представители семейства факторов Fox играют важную роль в этом сложном процессе, который окончательно не изучен.
У мышей некоторые гены, гомологичные гену caudal у дрозофил, экспрессируются исключительно в кишечнике. Cdx-1, экспрессирующийся в кишечнике взрослых организмов, также широко экспрессируется в тканях развивающегося эмбриона. Другой ген, Cdx-2, экспрессируется в висцеральной энтодерме на ранних стадиях эмбрионального развития, но впоследствии его паттерн экспрессии ограничен исключительно кишечником. Чрезмерная экспрессия Cdx-2 приводит к дальнейшей дифференцировке популяции клеток кишечника, которые в норме должны оставаться недифференцированными. В то же время эктопическая экспрессия Cdx-2 в стенке желудка приводит к формированию ткани, аналогичной кишечнику. Cdx-2 представляет собой важный фактор дифференцировки кишечника, однако до настоящего времени его роль в формировании кишечной трубки на ранних стадиях эмбрионального развития точно не установлена.