Формирование иммунитета у плода. Неонатальный лимфопоэз
Защиту организма от инфекционных агентов обеспечивают физические барьеры (кожа, слизистые оболочки и реснитчатый эпителий) и различные компоненты иммунной системы. Последняя состоит из Т- и В-лимфоцитов, NK-клеток, дендритных и фагоцитирующих клеток, а также белков комплемента. Иммунная система защищает организм также от аутоиммунных заболеваний и развития злокачественных опухолей.
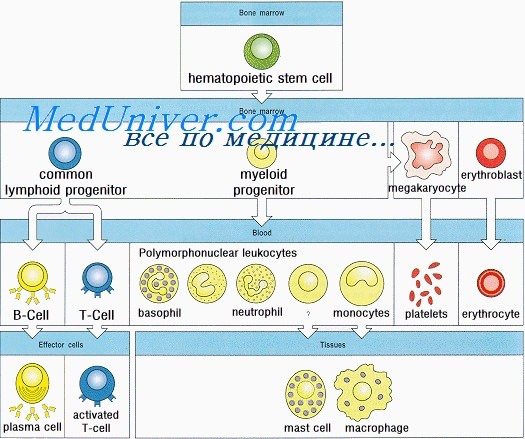
Иммунная система человека развивается из ткани первичной кишки. Полипотентные кроветворные стволовые клетки впервые появляются в желточном мешке на 2,5-3-й неделе внутриутробного развития и на 5-й неделе мигрируют в печень плода. Позднее они попадают в костный мозг, где и остаются на протяжении всей жизни. Из этих клеток образуются лимфоидные стволовые клетки, которые в зависимости от локализации дифференцируются в Т-, В- и NK-клетки. В середине I триместра беременности у плода образуются первичные лимфоидные органы (тимус и костный мозг), а вскоре и вторичные — селезенка, лимфатические узлы, миндалины, пейеровы бляшки и lamina propria. В этих органах дифференцировка Т-, В- и NK-лимфоцитов из стволовых клеток продолжается в течение всей жизни.
Как для органогенеза, так и для клеточной дифференцировки необходимо взаимодействие поверхностных молекул лимфоидных клеток с их микроокружением. Многочисленность и разнообразие этих поверхностных молекул отражена в международной классификации дифференцировочных антигенов, которые носят название кластеров дифференцировки (CD).
Т- и В-лимфоциты — единственные компоненты иммунной системы, способные распознавать специфические антигены; именно из них образуются клетки памяти (адаптивный иммунитет). Из кроветворных стволовых клеток образуются и NK-клетки, которые также являются лимфоцитами. Они защищают организм от вирусных инфекций, уничтожают опухолевые клетки и участвуют в регуляции иммунных реакций. Белки, синтезируемые и секретируемые Т- и В-лимфоцитами и NK-клетками, а также взаимодействующими с ними клетками, называются цитокинами. Некоторые из этих белков получили название интерлейкинов (ИЛ).
Цитокины играют роль ауто-, пара- и эндокринных факторов, способствующих дифференцировке и пролиферации клеток иммунной системы.
На 4-й неделе внутриутробной жизни из эктодермы третьей жаберной дуги и эндодермы третьего глоточного кармана образуется первичная закладка тимуса. На 7-8-й неделе правая и левая его части перемещаются каудально и сливаются друг с другом. На 8-й неделе предшественники Т-лимфоцитов из печени плода через кровь начинают поступать в окружающую тимус мезенхиму. Эти про-Т-лимфоциты идентифицируют по поверхностным белкам CD7 и CD34. К 8-8,5 нед. гестации СD7-клетки обнаруживаются уже в самом тимусе, причем на некоторых из них экспрессируются также CD4 и CD8.
Первый из этих маркеров характерен для зрелых Т-лимфоцитов-хелперов (Th), a CD8 — для зрелых цитотоксических Т-лимфоцитов и NK-клеток. Кроме того, на некоторых клетках присутствуют одиночные цепи антигенраспознающих рецепторов Т-лимфоцитов (TCR, Ti), но не полные TCR.
Зрелые TCR представляют собой гетеродимеры, состоящие из двух цепей — либо а и b, либо у и 8. TCR экспрессируются на клеточной поверхности вместе с CD3, образуя комплекс из пяти полипептидных цепей). Рекомбинация гена TCR осуществляется путем слияния крупных отдаленных друг от друга сегментов ДНК. Эти сегменты называются V (вариабельным), D (дополнительным) и J (соединяющим), причем каждый из них имеет ряд вариантов. Сегменты VDJ присоединяются к константной области а-гена, а сегменты VJ — к р-гену, завершая строение генов, кодирующих TCR. Случайная комбинация сегментов и определяет, в основном, огромное разнообразие TCR, придавая им способность распознавать миллионы различных антигенов. Рекомбинация генов TCR регулируется ферментами RAG-1 и RAG-2 (рекомбиназы, активирующие гены), а также другими ферментами.
При тяжелом комбинированном иммунодефиците у мышей и некоторых людей процесс рекомбинации нарушен. Рекомбинация генов TCR знаменует собой превращение про-Т-лимфоцитов в пре-Т-лимфоциты. Этот процесс начинается вскоре после заселения тимуса стволовыми клетками. На 8-10-й неделе внутриутробной жизни начинают формироваться Т-лимфоциты с разной функцией. На 9,5-10-й неделе более 95% тимоцитов экспрессирует CD7, CD2, CD4, CD8 и цитоплазматический CD3, а примерно 30% — внутренний антиген клеток коркового вещества тимуса. К 10-й неделе 25% тимоцитов обладают TCRaP. Число таких тимоцитов в ходе эмбрионального развития постепенно увеличивается и после рождения превышает 95 %.
По мере экспрессии TCR начинаются процессы позитивной и негативной селекции клеток коркового вещества тимуса. Позитивная селекция происходит путем взаимодействия незрелых тимоцитов (слабо экспрессирующих TCR) с МНС на эпителиальных клетках тимуса. В результате активируются и созревают только те тимоциты, рецепторы которых способны взаимодействовать с чужеродными антигенами, представляемыми им антигенами МНС. Зрелые тимоциты, прошедшие процесс селекции, экспрессируют либо CD4, либо CD8. Первые способны взаимодействовать с чужеродными антигенами, представляемыми молекулами HLA класса II, а вторые — с антигенами, представляемыми классом I HLA.
Затем начинается негативная селекция, обусловленная взаимодействием выживших тимоцитов, экспрессирующих значительно большее количество TCR, с пептидами хозяина, представляемыми HLA классов I и II, которые присутствуют на поступивших из костного мозга макрофагах, дендритных клетках и, возможно, В-лимфоцитах. Взаимодействие с этими антигенами включает программу апоптоза (запрограммированной гибели) аутореактивных тимоцитов. Клетки коркового вещества тимуса плода относятся к числу наиболее быстро делящихся клеток организма; их количество за 2 нед. после заселения тимуса стволовыми клетками возрастает в 100 000 раз. По мере их созревания происходят процессы селекции, в результате которых 97 % этих клеток погибает. Выжившие клетки несут на своей поверхности либо только CD4, либо только CD8, но не оба этих антигена и мигрируют в мозговое вещество тимуса.
Одновременно с появлением тимоцитов, обладающих только одним дифференцировочным антигеном, развиваются и функции Т-лимфоцитов, но до их выхода из тимуса эти клетки остаются незрелыми. Одна стволовая клетки дает начало примерно 3000 зрелых клеток мозгового вещества тимуса. Они устойчивы к лизирующему эффекту кортикостероидов. На 11—12-й неделе внутриутробной жизни Т-лимфоциты начинают покидать тимус, заселяя селезенку, лимфатические узлы и аппендикс, а на 14-15-й неделе — и миндалины.
Наибольшее их количество скапливается в корковом веществе лимфатических узлов, вокруг артериол селезенки и в лимфе грудного лимфатического протока. Клетки, недавно покинувшие тимус, одновременно экспрессируют изоформы CD45RA и CD62L (L-селектин). Рекомбинация генов TCR в ходе этого процесса приводит к образованию в качестве побочного продукта круговых эписом, которые обнаруживаются в Т-лимфоцитах, недавно покинувших тимус. В Т-лимфоцитах, созревающих вне тимуса, эписомы отсутствуют. Молекула адгезии, L-селектин, взаимодействует с углеводными компонентами специфических участков кровеносных сосудов. Такие участки (венулы с высоким эпителием) присутствуют в периферических лимфоидных органах, что и определяет задержку лимфоцитов в них. К 12-й неделе внутриутробной жизни Т-лимфоциты приобретают способность к пролиферации под действием растительных лектинов, таких как ФГА и КонА, и аллогенных клеток; антигенсвязывающие Т-лимфоциты появляются на 20-й неделе.
Тельца Гассаля (завитки полностью дифференцированных эпителиальных клеток мозгового вещества тимуса) впервые обнаруживаются на 16-18-й неделе внутриутробной жизни.