Патогенез пути NFKB/Rel в реакции эпителия кишечника на бактерии
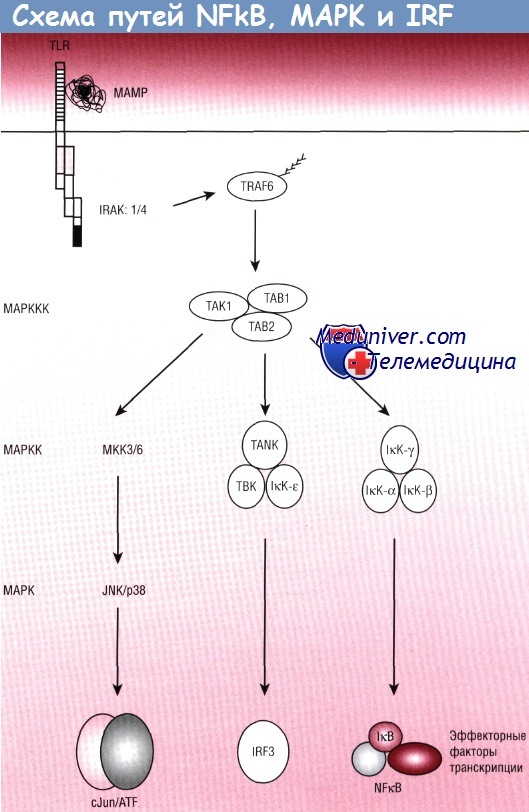
Присоединение PRR к соответствующим МАМР приводит к активации цитоплазматических сигнальных схем. К последним, в частности, относят классические пути NFKB/Rel, пути митоген-активируемой протеинкиназы (МАРК) и фактора транскрипции, индуцирующего интерфероновый ответ (IRF). Все три схемы используют посттрансляционные сигнальные реле, управляемые контролируемым и регулируемым трансфером ковалентных модификаций (фосфорилирования и убиквитинизации) вдоль ряда цитоплазматических белковых посредников. Активация путей NFkB, МАРК и IRF в итоге приводит к переносу в ядре факторов транскрипции семейств Rel, bZip и IRF соответственно.
При этом происходят присоединение специфических промоторных элементов и активация тех программ транскрипции генов, которые участвуют в защите организма хозяина.
NFkB в целом относится к членам семейства Rel ДНК-связывающих факторов транскрипции, которые связывают специфические последовательности мотивов в генных промоторах, регулирующих иммунный и воспалительный ответы. В кишечнике NFkB обычно существует в виде гетеродимера двух связанных белков — р65 и р50.
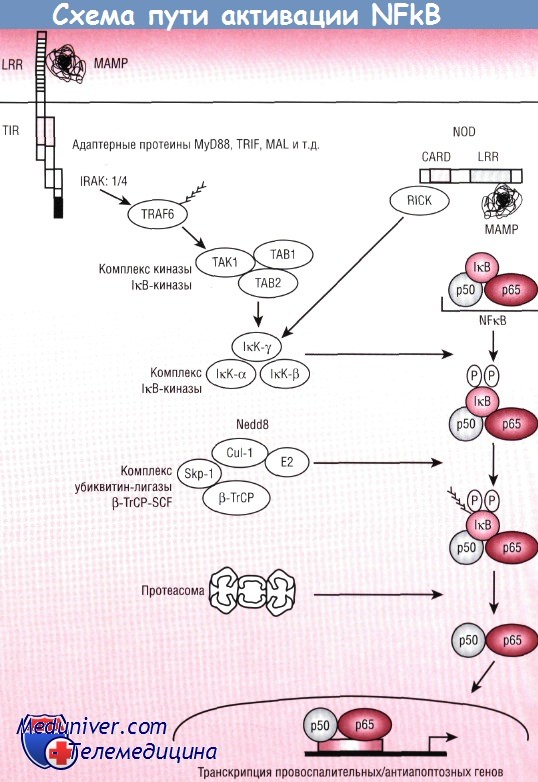
Активация NFkB тщательно регулируется, в норме процесс изолирован в цитоплазме 1 кВ. Сложная последовательность событий, в результате которой бактерии активируют NFkB посредством TLR и NOD-протеинов, схематично представлена на рисунке. Согласно существующей в настоящее время гипотезе, соответствующий МАМР присоединяется к TLR/NOD, что ведет к димеризации и образованию TIR-домена, который затем может присоединяться к классу переходных (адаптерных) белков.
Класс переходных белков состоит из MyD88, MAL/TIRAP и TRIF/TICAM. Появляются доказательства того, что эти переходные белки могут предпочтительно взаимодействовать со специфическими TLR и, предположительно, вовлекать большинство соответствующих сигнальных путей во взаимодействие между МАМР и TLR. Все адаптеры семейства MyD88 взаимодействуют со второй переходной молекулой киназы, ассоциированной с рецептором IL-1 (IRAK), некоторые члены семейства которой в настоящее время уже известны.
Трансмембранные TLR или интрацитоплазматические NOD-протеины связывают МАМР и передают сигналы с помощью цитоплазматических сигнальных посредников для активации IкВ-киназы. Этот комплекс фосфорилирует IкВ, который затем подвергается убиквитинации и распаду. Фосфорилирование обозначено на рисунке буквой Р, а убиквитинация изображена в виде «ершика».
CARD — домен активации и рекрутирования каспазы;
Е2 — убиквитинприсоединяющий фермент;
IкВ — ингибитор каппа В;
IкК— IкВ-киназа;
IRAK — киназа, ассоциированная с рецептором IL-1;
LRR — обогащенные лейцином повторы;
MAL — адаптерный белок, содержащий TIR-домен, подобный адаптеру MyD88;
МАМР — ассоциированные с микроорганизмами молекулярные паттерны;
MyD88 — белок 88 первичного ответа миелоидной дифференцировки;
NFkB — нуклеарный фактор каппа В;
NOD — нуклеотидсвязывающий олигомерный домен;
TIR — рецепторный домен Toll/IL-1;
TLR — Toll-подобные рецепторы;
TRAF6 — ассоциированный с TNF фактор 6;
TRIF — адаптерная молекула, содержащая адаптерный белок с TIR-доменом, индуцирующий IFN-b.
Кстати, специфическая роль данного белка в управлении бактериальным стрессом подтверждается наблюдением генетических нарушений в результате мутаций IRAK4, проявляющихся повторяющимися пиогенными инфекциями в детском возрасте. Затем IRAK, серинкиназа, активирует цитоплазматический сигнальный промежуточный ассоциированный с TNF фактор 6 (TRAF6), возможно посредством фосфорилирования. При активации TRAF6 становится убиквитинарным, что, в свою очередь, активирует киназу ТАК1 в комплексе с ТАВ1 и ТАВ2. Комплекс ТАК1/ТАВ1/ТАВ2 функционирует как киназа IкВ-киназы (IкКК), которая активирует комплексы IкВ-киназы (IкК).
1кК действует, вероятно, как сигнальное связующее звено, получающее и преобразующее сигналы от многих провоспалительных сигнальных трансдукционных путей. В дополнение к TLR-опосредованным сигналам многие стимулы активируют комплекс IкК, включая провоспалительные цитокины фактора некроза опухоли (TNF) и IL-1, которые через специфические рецепторы и сигнальные посредники действуют на IкК. После присоединения соответствующего МАМР к NOD-рецептору димеризация CARD индуцирует взаимодействие со вторым мессенджером — серин/треонинкиназой RICK, которая, в свою очередь, активирует 1кК и может напрямую активировать апоптоз.
Мобилизация Са2+ также может активировать эти пути. В каждом отдельном случае сигналы сходятся (конвергируют) на комплексе IкК. Основные каталитические составляющие комплекса, IкК-а и IкК-b, фосфорилируют два сериновых остатка с IкВ.
Фосфорилированный IкВ быстро становится полиубиквитинированным под действием комплекса убиквитин-лигазы (b-TrCP-SCF). Этот комплекс состоит из:
(1) b-TrСР, который физически взаимодействует с фосфо-IкВ;
(2) фермента убиквитин-лигазы (Е2);
(3) связующей субъединицы, названной Skp-1;
(4) регуляторной субъединицы Cul-1, которая сама по себе регулируется ковалентной модификацией убиквитин-подобной молекулы Nedd8.
В результате полиубиквитинации IкВ попадает в протеасому, что ведет к протеолитическому перевариванию молекулы IкВ. После распада IкВ NFkB перемещается вдоль ядерной мембраны с последующим присоединением ДНК к соответствующим промоторам, вызывая транскрипционную активность и синтез новой мРНК.
Было продемонстрировано, что взаимодействие непатогенных бактерий с моделью эпителия in vitro подавляло активацию NFkB на уровне убиквитинации IкВ. Впоследствии было показано, что это ингибирование происходит благодаря опосредованной бактериями потере NeddS-модификации регуляторной субъединицы Cul-1 и последующей потере ферментативной активности комплекса 1кВ/убиквитин-лигаза.
Эти открытия позволили высказать предположения о механизмах, посредством которых нормальная бактериальная колонизация в пищеварительном тракте может оказывать воздействие на воспалительные, определяющие в целом выживание организма пути.
В то время как NFkB, вероятно, является ключевым путем, активируемым микробными сигналами, два других пути могут быть быстро активированы TLR и, возможно, могут действовать как модификаторы для обеспечения специфических паттернов экспрессии, предназначенных для оптимального ответа при действии определенных классов микроорганизмов.