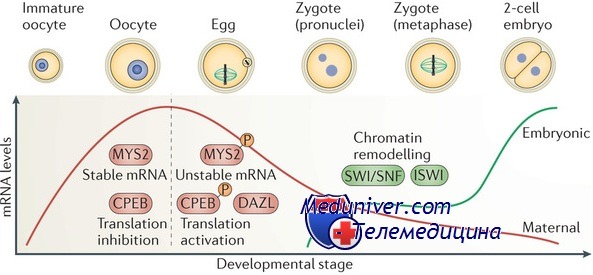
Кроме простого переноса материнского генома ооцит также несет с собой специфические генные продукты, необходимые для раннего эмбрионегеза вплоть до активации генома зиготы. У мышей активация генома зиготы происходит на двухклеточной стадии, у человека — на стадии 4-8 клеток.
Так как ооцит после возобновления мейоза не синтезирует новые транскрипты мРНК, а геном эмбриона не активируется раньше стадии 2 клеток, ооцит должен хранить транскрипты и белки, необходимые для его нужд в период ооцитарно-эмбрионального перехода. Гены, кодирующие белки ооцита, находящиеся в нем и выполняющие важнейшие функции на ранних стадиях эмбриогенеза, получили название «гены материнского эффекта».
Неудивительно, что несколько белков, кодируемых генами материнского эффекта [ооцит-специфичный линкерный гистон HI (H1F00), нуклеоплазмин 2 (Npm2), ооцитарная изоформа ДНК-метилтрансферазы-1 (Dnmtlo)], вовлечены в механизмы эпигенетической регуляции генов, которая начинается еще в ооците и продолжается в период раннего эмбриогенеза. Все эти известные гены материнского эффекта, а также те, которые еще не открыты, показывают, каким образом неоплодотворенный ооцит способен влиять на эмбрион, кроме своего вклада в собственно геном эмбриона.
H1F00 — ооцит-специфичный линкерный гистон H1, который связывается с хроматином на стадии ГП и дальнейшего созревания ооцита, вплоть до стадии двух-четырехклеточного эмбриона, когда он постепенно замещается соматическим H1. Так как соматические Н1-линкерные гистоны играют важную роль в регуляции функций хроматина, специфическая экспрессия H1F00 в ооцитах и ранних эмбрионах указывает на возможность его важной роли в ремоделировании хроматина при ооцитарно-эмбриональном переходе.
В отличие от H1F00, роль Npm2 (белка, содержащегося в ядрах ооцитов и эмбрионов в периимплантацион-ном периоде) в поддержании структуры хроматина уже доказана. У эмбрионов Npm2-/--самок мышей Npm2 отсутствует, и они погибают еще до имплантации, так как показано, что Npm2 необходим для поддержания нормальной структуры ядрышек. В частности, у них отсутствуют гетерохроматин и деацетилированный гистон НЗ, которые обычно располагаются в перинуклеолярном ободке ооцитов и ранних эмбрионов.
Соответственно, эта модель демонстрирует важную роль регуляции состояния хроматина и ядерной архитектоники для раннего эмбриогенеза.
Другой фактор, важный для эпигенетической регуляции геном материнского эффекта, — Dnmt1. Его ооцитарная форма (Dnmtlo) представляет собой изоформу белка Dnmtl, которая экспрессируется только в растущем ооците и раннем эмбрионе под воздействием ооцит-специфичного промотора. Эмбрионы, образовавшиеся из Dnmtlo-дефицитных ооцитов, также характеризуются отсутствием Dnmtlo и погибают на более поздних стадиях внутриутробного развития.
Эти эмбрионы характеризуются наличием дефектов импринтинга, о чем свидетельствуют неправильное деметилирование в определенных генных локусах и утрата аллель-специфичной экспрессии генов. Считают, что Dnmtlo особенно необходим на стадии 8 клеток, когда он временно перемещается из цитоплазмы, места постоянной локализации, в ядро.
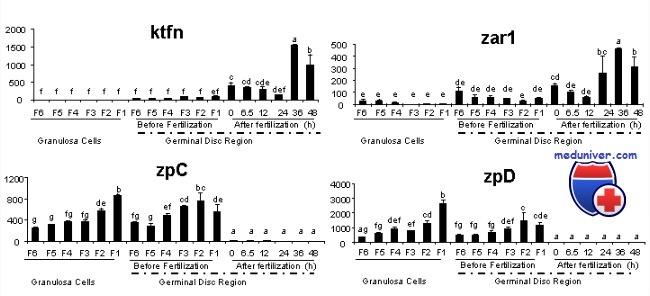
У самок мышей с инактивированным геном Zar1 имеет место нормальный оогенез, у них нормально запускается процесс оплодотворения, но пронуклеусы не сходятся и развитие большинства эмбрионов тормозится на стадии одной клетки. У тех единичных эмбрионов, которым удается дойти до двухклеточной стадии, наблюдают снижение экспрессии белков комплекса, необходимого для транскрипции, что свидетельствует о нарушении механизма активации эмбрионального генома.
В то время как Zar1 важен для развития эмбриона до двухклеточной стадии, Mater (другой белок, кодируемый геном материнского эффекта) экспрессируется в ооцитах и преимплантационном эмбрионе и играет огромную роль в развитии эмбриона на более поздней стадии. Изучение функций этих белков может приблизить нас к пониманию механизмов регуляции слияния отцовского и материнского геномов, активации эмбрионального генома и раннего эмбриогенеза.