Генетика контроля питания скелетных мышц. Влияние диеты на мышцы
Жизнеспособность клеток зависит от их возможности «добывать» питательные и другие необходимые вещества из окружающей клетки среды. Клетки синтезируют белки, предназначенные именно для этой цели. Если говорить в общих чертах, существуют две категории вовлеченных в этот процесс генов:
(1) гены, которые экспрессируют белки, транспортирующие нутриенты через мембраны клеток;
(2) гены, которые экспрессируют ферменты, задействованные в метаболизме нутриентов.
В обоих случаях гены регулируются для удовлетворения потребности какого-либо органа или системы. Эта форма регуляции генов в целом подобна таковой при регуляции генов у бактерий, т.е. триптофановому оперону. Клетки бактерий меняют свой фенотип для получения определенных преимуществ для себя, а не для удовлетворения каких-либо потребностей организма в целом.
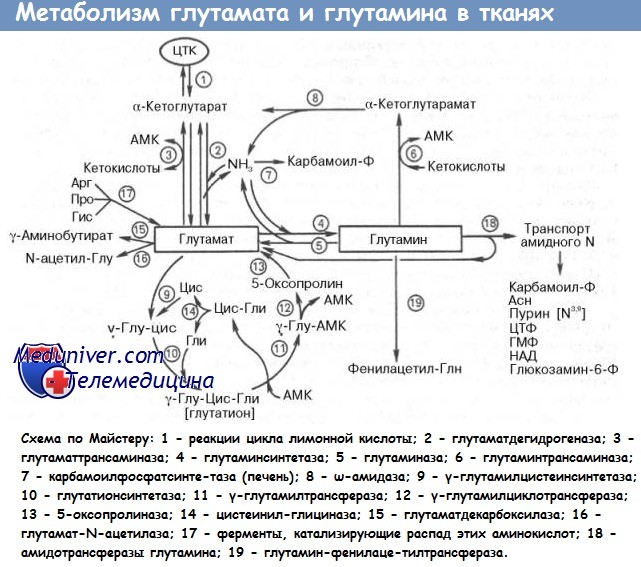
Примером того, как клетки могут с пользой для себя изменяться под воздействием внешней среды, может служить использование глютамина скелетными мышцами. Доступность глютамина влияет на процессы обмена в мышцах. Кроме того, глютамин является важнейшим источником аминоазота для мышечных клеток. Глютамин также играет значимую роль в регуляции быстро делящихся клеток, таких как лимфоциты и эпителиальные клетки кишечника.
Глютамин и глютамат транспортируются в клетку с помощью натриевых переносчиков. Для экспериментальных целей в качестве первичной клеточной культуры может быть взята скелетная мышца крысы. Эти клетки демонстрируют повышение экспрессии внутреннего транспорта как глютамина, так и глютамата, когда они лишены экзогенного обеспечения глютамином, к тому же одновременно увеличивается активность глютаминсинтетазы.
Этот фермент катализирует присоединение аминогруппы к карбоксильной части молекулы глютамата, в результате образуется глютамин. Таким образом, два различных нутриента, глютамин и глютамат, могут полностью обеспечить потребность мышечных клеток в глютамине. Это обеспечение достигается двумя независимыми путями, каждый из которых находится под генетическим контролем.
Описанный механизм демонстрирует, как различные нутриенты могут перекрестно стимулировать экспрессию связанных между собой белков: удаление глютамина или глютамата из окружения клетки ведет к усилению транспорта обеих аминокислот. Не только дефицит глютамина и глютамата усиливает экспрессию их собственных транспортных белков, но и утрата одного вещества приводит к повышению экспрессии переносчика другого.
Исследование кинетики транспорта показало, что дефицит глютамина (или глютамата) усиливал максимальную скорость, с которой аминокислота транспортировалась в клетку (Vmax), и не изменял аффинность переносчика к соответствующей аминокислоте. Это позволило предположить, что возникающий дефицит аминокислот приводит к увеличению продукции ряда соответствующих переносчиков. Время, необходимое для удвоения Vmax, составляло примерно 4 час. Данный факт, а также то, что индукция обоих переносчиков (как глютамата, так и глютамина) прекратилась, когда глютамин был удален в присутствии актиномицина D (который ингибирует синтез ДНК), свидетельствует о том, что их регуляция осуществляется посредством инициации транскрипции.
В транспортных системах глютамина и глютамата еще многое непонятно, но исследования в этой области весьма перспективны, т.к. каждая аминокислота имеет только два переносчика — натрий-зависимый и натрий-независимый. Только натрий-зависимый переносчик реагирует на исчезновение какого-либо нутриента. Регуляция глютамата и глютамина, несмотря на свою важность, составляет лишь малую часть нутриентного гомеостаза в клетках скелетных мышц in vivo, поскольку их энергетические потребности в основном удовлетворяются посредством глюкозы и свободных жирных кислот.
До сих пор неясно, до какой степени изменения в диете влияют на способность скелетной мышцы регулировать потребление жиров. Cameron-Smith и соавт. показали, что краткосрочная диета с высоким содержанием жиров повышает экспрессию генов липидного метаболизма и экспрессию генов в мышце человека. Они обследовали 14 спортсменов, которых разделили на группы.
Одна группа находилась на диете с высоким содержанием жиров, а другая — на изоэнергетической диете с высоким содержанием углеводов. Диету применяли в течение 5 дней с 2-недельным перерывом и последующей перекрестной сменой диет. При этом спортсмены получали определенную физическую нагрузку. В первый день и после завершения курса каждой из диет у обследованных была выполнена биопсия мышц и взяты анализы крови. По сравнению с диетой с большим содержанием углеводов «жировая» диета обусловливала более выраженную генную экспрессию транслоказы жирных кислот (предполагаемого переносчика жирной кислоты) и b-гидроксиацил-СоА-дегидрогеназы (фермента b-окисления), а также большее количество транслоказы жирных кислот.
Это подразумевает повышение экспрессии генов, необходимых для транспортировки жирных кислот и окислительного метаболизма в скелетной мышце. По заключению исследователей взаимодействие жирной кислоты с генетическим материалом скелетных мышц представляется важным из-за вклада этого процесса в способность окислительного профиля мышцы к адаптации к доминирующему источнику пищи. Однако необходимо более точное объяснение этих механизмов.