Генетическая регуляция дифференцировки стволовых клеток тонкой кишки
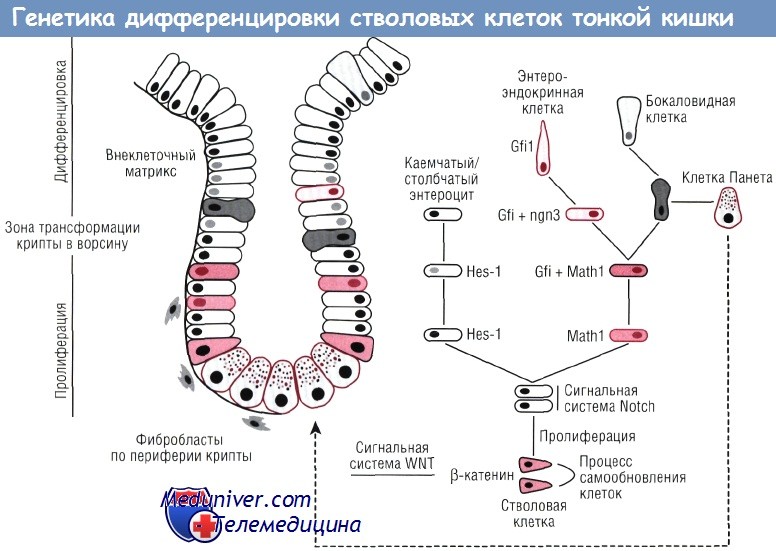
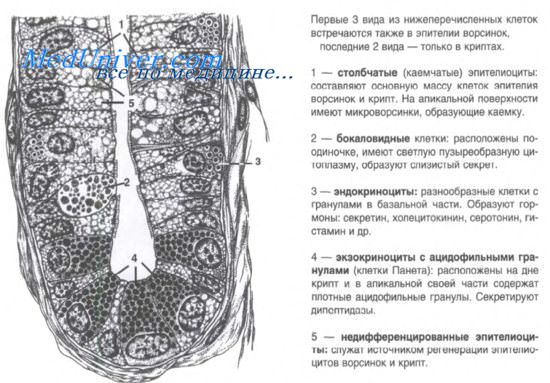
Из стволовых клеток происходят четыре популяции клеток тонкой кишки. При использовании различных подходов установлено, что ключевую роль в дифференцировке отдельных клеточных популяций играет сигнальный путь Notch. Сверхэкспрессия Notch сопровождается исчезновением бокаловидных, энтероэндокринных клеток и клеток Панета при одновременном увеличении популяции пролиферирующих клеток. Сигнальный путь Notch активирует Hes-1 и подавляет факторы Math1 и ngn3, что подтверждает его роль в дифференцировке клеток.
Изменение сигнального пути Notch за счет выключения промежуточных ключевых звеньев этого пути сопровождается гиперплазией бокаловидных клеток путем пролиферации клеток-предшественников. Аналогично применение растворимых ингибиторов у-секретазы, тормозящих активацию сигнального пути Notch, сочетается с увеличением количества бокаловидных клеток в тонкой кишке. Кроме того, обнаружено, что в развивающейся кишке под воздействием факторов Notch происходит обратимая остановка процесса морфогенеза, сопровождающаяся существенным уменьшением количества клеток-предшественников.
В то же время во взрослом организме факторы Notch оказывают воздействие на клетки, не затрагивая процесс морфогенеза. Установлена также роль таких факторов, как Math1, Hes-1 и ngn3, которые являются мишенями сигнального пути Notch. Выключение Math1, фактора транскрипции, играющего большую роль в дифференцировке нервных клеток, сопровождается исчезновением клеток Панета, бокаловидных и энтероэндокринных клеток, при этом сохраняются только лишь каемчатые или столбчатые энтероциты.
Таким образом, Mathl представляет собой основной фактор, необходимый для формирования популяции секреторных клеток. Предполагают, что Mathl подавляется экспрессией другого фактора транскрипции — Hes-1, который исчезает в популяции секреторных клеток, но продолжает вырабатываться на ранних стадиях развития каемчатых энтероцитов. У лабораторных мышей при нокауте гена Hes-1 обнаружено увеличение количества эндокринных клеток как в кишке, так и в поджелудочной железе. Данный факт свидетельствует о том, что Hes-1 представляет собой ингибитор популяции эндокринных клеток. Клетки-предшественники секреторной популяции изначально вырабатывают как Math1, так и фактор транскрипции Gfi1.
Впоследствии происходит формирование двух клеточных популяций — энтероэндокринных клеток, экспрессирующих Gfi1 и ngn3, и клеточной линии, из которой в дальнейшем образуются клетки Панета и бокаловидные клетки, продуцирующие Math1 при одновременной блокировке продукции Gfi1. Таким образом, в соответствии с современными представлениями сигнальный путь Notch, действуя между смежными клетками, увеличивает экспрессию фактора Hes. В свою очередь, увеличение фактора Hes сопровождается подавлением экспрессии Math1, что приводит к трансформации клеток в каемчатые энтероциты. В клетках с низкой экспрессией Hes усиливается продукция Math1, результатом чего является формирование популяции секреторных клеток.
Энтероэндокринные клетки характеризуются экспрессией Gfi1, но продукция Gfi1 прекращается в клеточной популяции, дающей начало клеткам Панета и бокаловидным клеткам. Фактор Hes-1 также необходим для образования энтероэндокринных клеток желудка и поджелудочной железы, как и для образования энтероэндокринных клеток тонкой кишки. Фактор KLF4 нужен для формирования бокаловидных клеток толстой кишки, но его присутствие не требуется в процессе развития аналогичной клеточной популяции тонкой кишки. В противоположность селективной аблации популяции клеток желудка удаление клеток Панета в тонкой кишке не оказывает влияния на другие клеточные линии тонкой кишки.
В одном из недавно опубликованных, представляющих интерес наблюдений приводится описание больного с тяжелым энтероколитом, сопровождающимся достоверно установленным отсутствием клеток Панета, бокаловидных и энтероэндокринных клеток. Указанное наблюдение имеет поразительное сходство с моделью трансгенных мышей с делецией фактора Math1, у которых также отсутствовали все три указанные клеточные популяции. Молекулярная основа дефекта, обнаруженного у пациента, не была установлена.
Следствием унитарной гипотезы происхождения эпителиальных клеток стало предположение, что крипты содержат промежуточные клетки, которые не окончательно коммитированы в одну конкретную клеточную популяцию. Нейрогенин-3 необходим для процесса дифференцировки эндокринных клеток кишки. У мышей с нокаутом ngn3 в половых клетках не происходит формирования эндокринных клеток в поджелудочной железе или кишке, но по-прежнему образуются эндокринные клетки желудка. У данных мышей экспрессируется Mathl. Это дополнительно свидетельствует о том, что Math1 влияет на ngn3. В сформированной кишке ngn3 вырабатывается в небольшом количестве клеток, расположенных вблизи основания крипты, что свидетельствует о продукции данного фактора на ранних стадиях дифференцировки.
По-видимому, ngn3 вырабатывается в стволовых клетках либо в их потомках — промежуточных клетках-предшественниках.
На основании анализа клеточных популяций установлено, что клетки-предшественники дифференцируются как в эндокринные, так и в не обладающие подобной функцией клеточные линии. Указанный факт свидетельствует о том, что ngn3 первоначально экспрессируется в клетках-предшественниках, которые еще не вступили в необратимый процесс дифференцировки в эндокринные клетки.
Блок клеточного цикла тесно связан с окончательной дифференцировкой эпителиальных клеток кишки, однако механизм данного процесса остается пока неизученным. Выработка с-тус контролируется сигнальной системой WNT и представляет собой основной фактор в процессе перехода от пролиферации к дифференцировке. Кроме того, установлено, что система с-тус активируется в клетках-предшественниках кишки. В то же время делеция гена с-тус сопровождается временным угнетением формирования крипт, но не оказывает существенного влияния на последующие процессы развития. В исследованиях in vitro установлено значение двух ингибиторов клеточной пролиферации — р21 и р27, однако их селективная делеция не сопровождалась изменениями фенотипа кишки in vivo. Вероятно, имеет место тесное взаимодействие многочисленных факторов различных семейств, и данный процесс остается предметом дальнейших исследований. Поиск механизмов регуляции развития кишечника усложняется тем фактом, что дифференцировка каемчатых энтероцитов и клеток Панета может индуцироваться фактором Rac1 без блокирования процессов пролиферации.
В лаборатории, возглавляемой Gordon, изучена экспрессия генов в клетках-предшественниках, изолированных с помощью лазерных скальпеля и пинцета. В качестве модели были использованы лабораторные мыши, у которых удаляли клетки Панета с целью компенсаторного увеличения количества потенциальных стволовых клеток. При попытке установить гены, оказывающие наиболее важное влияние на функцию стволовых клеток, было обнаружено большое количество генов с повышенной или пониженной экспрессией. Аналогичное исследование было выполнено в отношении клеток-предшественников желудка. При исследовании эпителиальных стволовых клеток как тонкой кишки, так и желудка обнаружено несколько молекулярных механизмов и вариантов паттернов экспрессии генов.
При сравнении данных паттернов с паттернами экспрессии, установленными при исследовании гемопоэтических стволовых клеток, не удалось обнаружить каких-либо общих генов. Это может отражать объективные сложности, возникающие при сравнении результатов различных исследований, либо свидетельствовать о том, что стволовые клетки в различных тканях обладают индивидуальными уникальными особенностями. Остается неясным, существует ли некоторый набор универсальных «стволовых» генов либо имеются уникальные варианты экспрессии генов, специфических для стволовых клеток в каждой конкретной ткани.