а) Секреция протонов и реабсорбция ионов бикарбоната в почечных канальцах. Секреция протонов и реабсорбция ионов бикарбоната происходят практически во всех частях канальцевой системы, за исключением тонкого отдела петли Генле. В целом процесс реабсорбции бикарбоната по ходу тубулярного сегмента нефрона изображен на рисунке ниже.
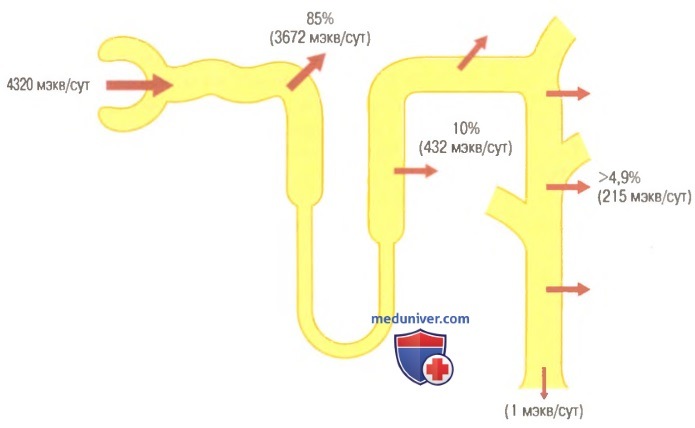
Реабсорбция бикарбонатов в различных отделах канальцевой системы нефрона, выраженная в процентах и миллиэквивалентах. Приведенные значения соответствуют нормальной реабсорбции за сутки
Следует помнить, что для реабсорбции каждого иона бикарбоната в просвет канальца необходимо секретировать один протон.
Около 80-90% реабсорбции ионов бикарбоната (и канальцевой секреции протонов) приходится на проксимальный каналец, поэтому лишь небольшое их количество поступает в дистальные сегменты нефрона. В толстом восходящем отделе петли Генле реабсорбируются другие 10% бикарбоната, прошедшего через почечный фильтр, а остаток поглощается дистальным канальцем и собирательной трубочкой. Как уже обсуждалось ранее, механизм реабсорбции бикарбонатов связан с канальцевой секрецией протонов, однако в различных сегментах канальцевой системы этот процесс протекает по-разному.
б) Секреция ионов водорода в проксимальных сегментах нефрона осуществляется благодаря вторично активному транспорту. Эпителиальные клетки проксимального канальца, толстого восходящего отдела петли Генле, начальных отделов дистальных канальцев секретируют протоны в просвет канальцев благодаря контртранспорту с ионами Na+ (для облегчения понимания просим вас изучить рисунок ниже).
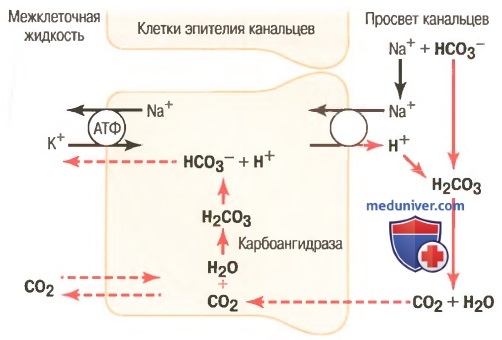
Внутриклеточные механизмы: (1) активная секреция протонов в просвет канальцев; (2) реабсорбция ионов бикарбоната в результате химического взаимодействия с протонами, образования угольной кислоты, которая разлагается на углекислый газ и воду; (3) реабсорбция ионов Na+ в обмен на протоны. Такой характер секреции ионов Н+ характерен для проксимальных канальцев, толстого восходящего отдела петли Генле и начальных отделов дистальных канальцев
Этот вторично активный механизм секреции связан с переносом ионов Na+ в клетку через апикальную мембрану с помощью белка, обменивающего натрий на протон.
Источником энергии для перемещения ионов Н+ в клетку служит производная градиента концентрации для Na+, который создается работой Na+/K+-насоса (АТФ-азы), расположенной на базолатеральной мембране эпителия канальцев. Таким способом реабсорбируется более 90% бикарбоната, что требует секреции в просвет канальцев ионов Н+ около 3900 мэкв/сут. Тем не менее, этот механизм не обеспечивает создание высокой концентрации ионов Н+ в моче: жидкость в просвете канальцев закисляется лишь в собирательных трубочках и каналах.
На рисунке выше показано, как при помощи процесса секреции ионов Н+ происходит реабсорбция бикарбоната. Секреция протонов начинается, когда СО2, попавший в клетку эпителия путем диффузии или в результате метаболических процессов, под влиянием фермента карбоангидразы соединяется с молекулой воды, формируя угольную кислоту. Угольная кислота диссоциирует на ионы Н+ и HCO3-.
Протоны выделяются в просвет канальца с помощью противотранспорта с ионами Na+. Другими словами, перед тем как ионы Na+ переместятся через апикальную мембрану внутрь клетки, они вначале объединяются с мембранным белком-переносчиком. В то же время с другой стороны к белку-переносчику присоединяется ион Н+. Ион Na+ перемещается в клетку по градиенту концентрации, установленному АТФ-азой Na+/K+-насоса, расположенного на базолатеральной поверхности клетки. Энергия, затраченная на создание градиента концентрации, обеспечивает движение ионов Na+ внутрь клетки, а также обусловливает перемещение ионов Н+ в противоположном направлении: из клетки в просвет канальца.
Ионы HCO3-, образованные в клетке при диссоциации молекулы H2CO3, затем пассивно перемещаются через базолатеральную мембрану в межклеточную жидкость почечной ткани и оттуда — через перитубулярные капилляры в кровь. Таким образом, на каждый выделенный в просвет канальца ион Н+ приходится один ион бикарбоната, реабсорбированный в кровь.