Как и в скелетной мышце, пусковым стимулом для сокращения большинства гладких мышц является увеличение количества внутриклеточных ионов кальция. В разных типах гладких мышц это увеличение может быть вызвано нервной стимуляцией, гормональной стимуляцией, растяжением волокна или даже изменением химического состава окружающей волокно среды.
Однако в гладких мышцах нет тропонина (регуляторного белка, который активируется кальцием). Сокращение гладкой мышцы активируется совершенно другим механизмом, изложенным далее.
Соединение ионов кальция с кальмодулином. Активация миозинкиназы и фосфорилирование головки миозина.
Вместо тропонина гладкомышечные клетки содержат большое количество другого регуляторного белка, называемого кальмодулином. Хотя этот белок похож на тропонин, он отличается способом запуска сокращения. Кальмодулин делает это путем активации миозиновых поперечных мостиков. Активация и сокращение осуществляются в следующей последовательности.
1. Ионы кальция связываются с кальмодулином.
2. Комплекс кальмодулин-кальций соединяется с фосфорилирующим ферментом миозинкиназой и активирует ее.
3. Одна из легких цепочек каждой головки миозина, называемая регуляторной цепочкой фосфорилируется под действием миозинкиназы. Когда эта цепочка не фосфорилирована, циклического прикрепления и отделения миозиновой головки по отношению к актиновой нити не происходит. Но при фосфорилировании регуляторной цепочки головка приобретает способность к повторному связыванию с актиновой нитью и осуществлению всего циклического процесса периодических «подтягиваний», лежащих в основе сокращения, как и в скелетной мышце.
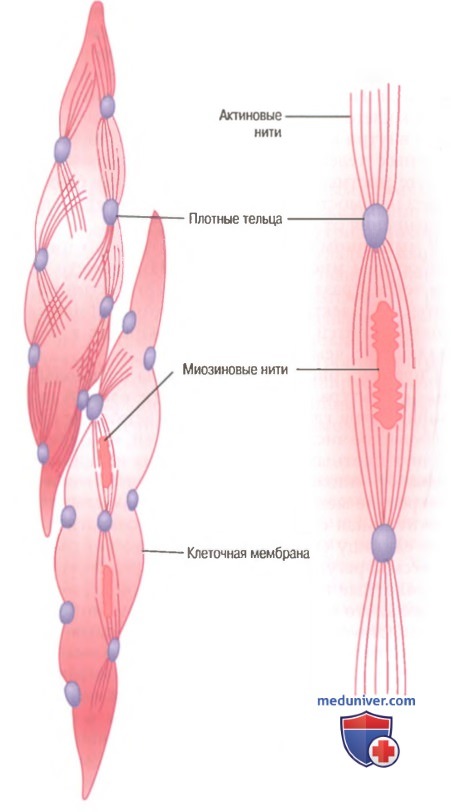
Морфологическая структура гладкой мышцы. В волокне слева вверху видны актиновые нити, исходящие из плотных телец. В волокнах слева внизу и справа на рисунке показано взаимоотношение между миозиновыми и актиновыми нитями.
Прекращение сокращения. Роль миозинфосфатазы. Когда концентрация ионов кальция падает ниже критического уровня, изложенные процессы автоматически развиваются в обратном направлении, кроме фосфорилирования головки миозина. Для обратного развития этого состояния нужен другой фермент — миозинфосфатаза, который локализуется в жидкостях гладкомышечной клетки и отщепляет фосфатазу от регуляторной легкой цепочки. После этого циклическая активность, а значит и сокращение, прекращается.
Следовательно, время, необходимое для расслабления мышцы, в большой степени определяется количеством активной миозинфосфатазы в клетке.
Возможный механизм регуляции механизма «защелки». В связи с важностью механизма «защелки» в функции гладких мышц предпринимаются попытки объяснить это явление, поскольку оно делает возможным долговременное поддержание тонуса гладких мышц многих органов без значительных энергетических затрат. Среди многих предложенных механизмов приводим один из простейших.
Когда сильно активированы и миозинкиназа, и миозинфосфатаза, частота циклов миозино-вых головок и скорость сокращения высокие. Затем, когда активация ферментов снижается, частота циклов уменьшается, но в то же время деактивация этих ферментов позволяет миози-новым головкам оставаться прикрепленными к актиновым нитям в течение все более длительной части цикла. Следовательно, число головок, прикрепленных к актиновой нити в любой данный момент времени, остается большим.
Поскольку число прикрепленных к актину головок определяет статическую силу сокращения, напряжение удерживается, или «защелкивается». Однако энергии при этом используется мало, поскольку расщепления АТФ до АДФ не происходит, за исключением тех редких случаев, когда какая-нибудь головка отсоединяется.
Видео физиология мышц и мышечного сокращения - профессор, д.м.н. П.Е. Умрюхин