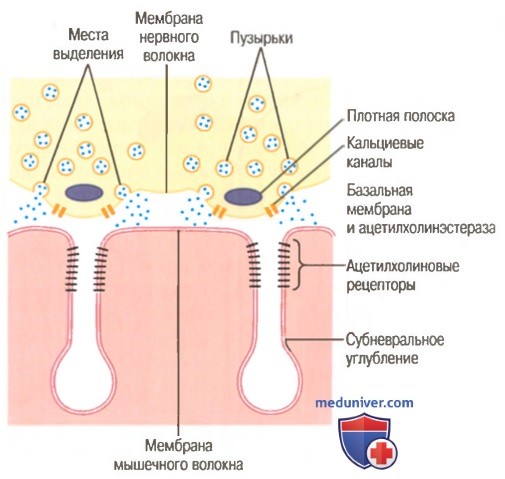
Механизм обмена ацетилхолина. Лекарственные средства влияющие на нервно-мышечное соединение
В связи с достаточно большим размером нервно-мышечного соединения оно является одним из немногих синапсов нервной системы, в котором раскрыты практически все детали химического проведения.
Так, известны следующие фазы процесса образования и выделения ацетилхолина в этом соединении.
1. Маленькие везикулы размером около 40 нм формируются в аппарате Гольджи в теле мотонейрона спинного мозга. Затем эти везикулы транспортируются аксоплазмой, которая «течет» внутри аксона на всем его протяжении от тела клетки в спинном мозге до периферических нервных окончаний аксона в нервно-мышечном соединении. Около 300000 этих маленьких пузырьков собираются в нервных окончаниях одиночной концевой пластинки в скелетной мышце.
2. Ацетилхолин синтезируется в цитозоле окончания нервного волокна и сразу же транспортируется через мембраны везикул внутрь, где он хранится в высокой концентрации (около 10000 молекул ацетилхолина в каждой везикуле).
3. Прибытие потенциала действия в нервное окончание открывает большое количество электроуправляемых кальциевых каналов в мембране нервного окончания. В результате концентрация ионов кальция с внутренней стороны мембраны окончания повышается более чем в 100 раз, что, в свою очередь, увеличивает скорость слияния везикул ацетилхолина с мембраной окончания примерно в 10000 раз. Это слияние приводит к разрыву множества везикул и делает возможным экзоцитоз ацетилхолина в синаптическое пространство. В ответ на каждый потенциал действия обычно разрываются около 125 везикул. Через несколько миллисекунд ацетилхолин расщепляется ацетилхолинэстеразой на ион ацетата и холин, который активно реабсорбируется внутрь нервного окончания, где используется для формирования нового ацетилхолина. Эта последовательность событий происходит в течение 5-10 мсек.
4. В нервном окончании количества доступных для использования везикул достаточно для передачи лишь нескольких тысяч импульсов с нерва на мышцу. Следовательно, для непрерывного функционирования нервно-мышечного соединения необходимо быстрое образование новых везикул. В течение нескольких секунд после окончания каждого потенциала действия в мембране нервного окончания появляются окаймленные ямки. Их появление связано с функцией сократительных белков нервного окончания, особенно белка клатрина, который прикрепляется к мембране в области образования везикул. В течение примерно 20 сек белки сокращаются, вызывая отрыв ямок от внутренней стороны мембраны с формированием новых везикул. Через несколько секунд внутрь этих везикул транспортируется ацетилхолин, и везикулы готовы для нового цикла выделения ацетилхолина.
Выделение ацетилхолина из синаптических пузырьков в пресинаптической мембране нервно-мышечного соединения.
Обратите внимание на близость участков освобождения медиатора в пресинаптической мембране к ацетилхолиновым рецепторам мембраны мышечного волокна в устьях субневральных углублений.
Лекарственные средства, усиливающие или блокирующие передачу в нервно-мышечном соединении
Лекарственные средства, стимулирующие мышечное волокно подобно ацетилхолину. Многие химические соединения, включая метахолину карбахол и никотин, оказывают такое же влияние на мышечное волокно, как и ацетилхолин. Различие между действием этих лекарственных веществ и ацетилхолином заключается в том, что лекарства не расщепляются холинэстеразой или разрушаются так медленно, что их действие часто продолжается от нескольких минут до нескольких часов. Эти лекарственные вещества вызывают локальную деполяризацию мембраны мышечного волокна в области двигательной концевой пластинки, где располагаются рецепторы для ацетилхолина. В результате каждый раз после восстановления мышечного волокна от предыдущего сокращения эти деполяризованные за счет утечки ионов области инициируют новый потенциал действия, вызывая состояние мышечного спазма.
Лекарственные средства, стимулирующие нервно-мышечное соединение путем инактивации ацетилхолинэстеразы. Хорошо известны три лекарственных средства (неостигмину физостигмин и диизопропилфторфосфат), которые инактивируют ацетилхолинэстеразу в синапсах, в результате гидролиз ацетилхолина не происходит. С каждым последующим нервным импульсом накапливается дополнительный ацетилхолин, повторно стимулирующий мышечное волокно. Это вызывает мышечный спазм даже при незначительном числе нервных импульсов, поступающих к мышце. К сожалению, это может привести к смерти за счет удушья из-за ларингоспазма.
Неостигмин и физостигмин, соединяясь с ацетилхолинэстеразой, могут тормозить ее действие в течение нескольких часов, затем данные вещества отделяются от ацетилхолинэстеразы, и она вновь становится активной. Напротив, диизопропилфторфосфат, который является боевым отравляющим нервно-паралитическим газом, инактивирует ацетилхолинэстеразу в течение недель, что делает этот газ смертельно опасным.
Лекарственные вещества, блокирующие передачу в нервно-мышечном соединении. Группа лекарственных веществ, известных как курареподобные вещества, могут предупреждать проведение импульсов с нервного окончания на мышцу. Например, D-тубокурарин блокирует действие ацетилхолина на ацетилхолиновые рецепторы мышечного волокна, предупреждая повышение проницаемости каналов мышечной мембраны до уровня, достаточного для возникновения потенциала действия.